



ПИШЕ: Милан Радуловић ([email protected])
Објављено: 30.11.2020. | Допуњено: 30.11.2020.
Преузмите бесплатно PDF верзију чланка.
ОПШТЕ НАПОМЕНЕ
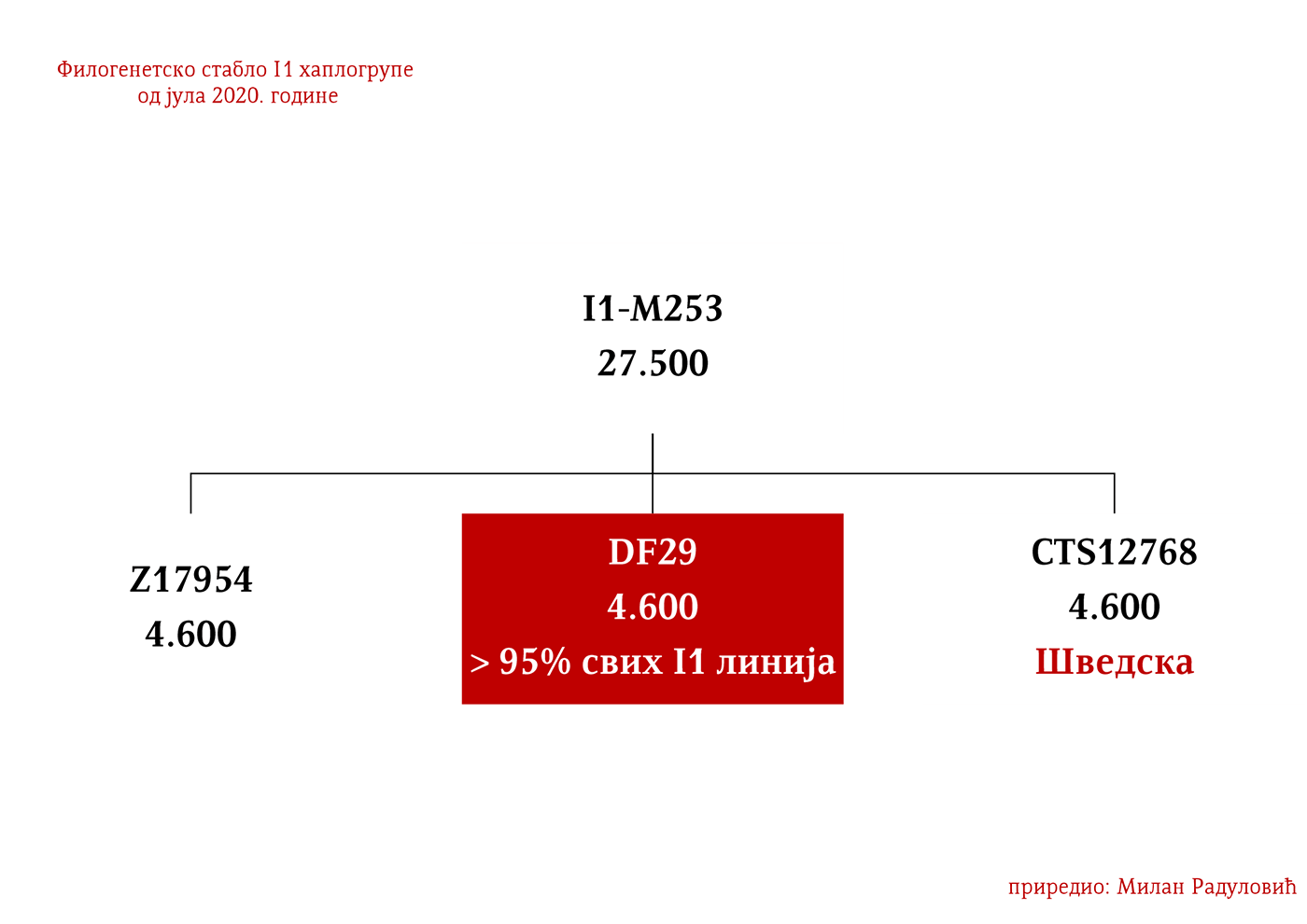
Хаплогрупа I1 проистекла је из предачке хаплогрупе I. Она се од ње одвојила прије 27 500 година. Заједнички предак свих данашњих припадника хаплогрупе I1 живио је прије 4 600 година и то негдје на простору централне Европе. Хаплогрупа I1 може се повезати са експанзијом германских народа. Према подацима Српског ДНК пројекта, она је код Срба заступљена у проценту од 8,32%.
ПОРИЈЕКЛО И МИГРАЦИЈЕ
Хаплогрупа I1 настала је прије 27 500 година и била је на ивици опстанка до прије 4 600 година када је живио како је претходно поменуто заједнички предак свих данашњих I1 мушкараца (YFull, 2020), могуће на простору Данске (Nordtvedt, 2008). За мјесто настанка хаплогрупе сматра се шири простор Европе. Представљали су прве ловце сакупљаче Европе заједно са припадницима I2 хаплогрупе, а у неолит су ушли са Културом линеарнe керамике у којој су били активни судионици. Најстарији узорци су из Шпаније и Шведске. У Шпанији су пронађени узорци који су предачки у односу на I1-M253 мутацију, BAL001/005 из горњег паолелита датиран у 11 384-10 733 п.н.е. са локалитета Балма Гилања у Каталонији (Villalba-Mouco, et al., 2019) и I10899/Car1 датиран у 9700–5500. год. п.н.е. (Olalde, et al., 2019). Ширење по Европи започињу током гвозденог доба и интензивно током антике и раног средњег вијека.
МЕЗОЛИТ
Током мезолита, ловачко-сакупљачког периода, I1 мушкарци били су распрострањени од Пиринејског полуострва до централне Европе, на шта нам указују налази древне ДНК. Када су први људи крочили на тло Скандинавије после повлачења леда прије неких 11 700 година, формирали су и прве културе. На маленом острву Карсле, близу Готланда, на локалитету Стура Карлсе, међу ловачко-сакупљачком заједницом пронађен је узорак који претходи I1 хаплогрупи код анализираног мушког скелета SF11 датираног у период између 7050-6850. год. п.н.е. (Günther, et al., 2018); док остала три скелета из истог рада припадају I2 хаплогрупи, односно узорак SBj.SG из Готланда датиран између 7050-6650. год. п.н.е. хаплогрупе I2a > Y4213; други скелет под шифром Steigen.SG из Молеја, Стејгена, Норвешка, датиран је између 4050-3850. год. п.н.е., I2a > M423; и трећи I2c > S6635, Hum2.SG, 7550-7350. год. п.н.е., такође из Норвешке, са локалитета Хумервиколмен. Оно што је уочљиво из рада јесте да тестирани узорци са истока Скандинавије имају претежно западне генетичке примјесе ловачко-сакупљачких заједница, док утицај источноевропских ловачко-сакупљачких заједница уочавамо у западним дјеловима Скандинавије. Правци миграција источних и западних ловачко-сакупљачких заједница као и њихови удјели у мезолитском становништву Скандинавије приказани су у Прилогу 1. Налаз SF11 у самом раду није означен као претходник I1 хаплогрупе, због лошије покривености Y-ДНК, па је овом приликом приступљено анализи сировог генетичког материјала ради потврде писања Генетикера (Genetiker, 2018) који је уочио да се ради о мушкарцу са мутацијом која је предачка у односу на I1 хаплогрупу. Након обраде сирових података, потврђујемо да се ради о мушкарцу са мутацијама које су рођачке у односу на I1-M253, а то су следеће: Z2726 G+, YSC0000301/Z2882 C+, CTS6221 T+, CTS1748 G+, Z2731 A+, CTS6629 A+, CTS5887 C+, Y7398 T+, док снип M253 није покривен лабораторијском анализом.
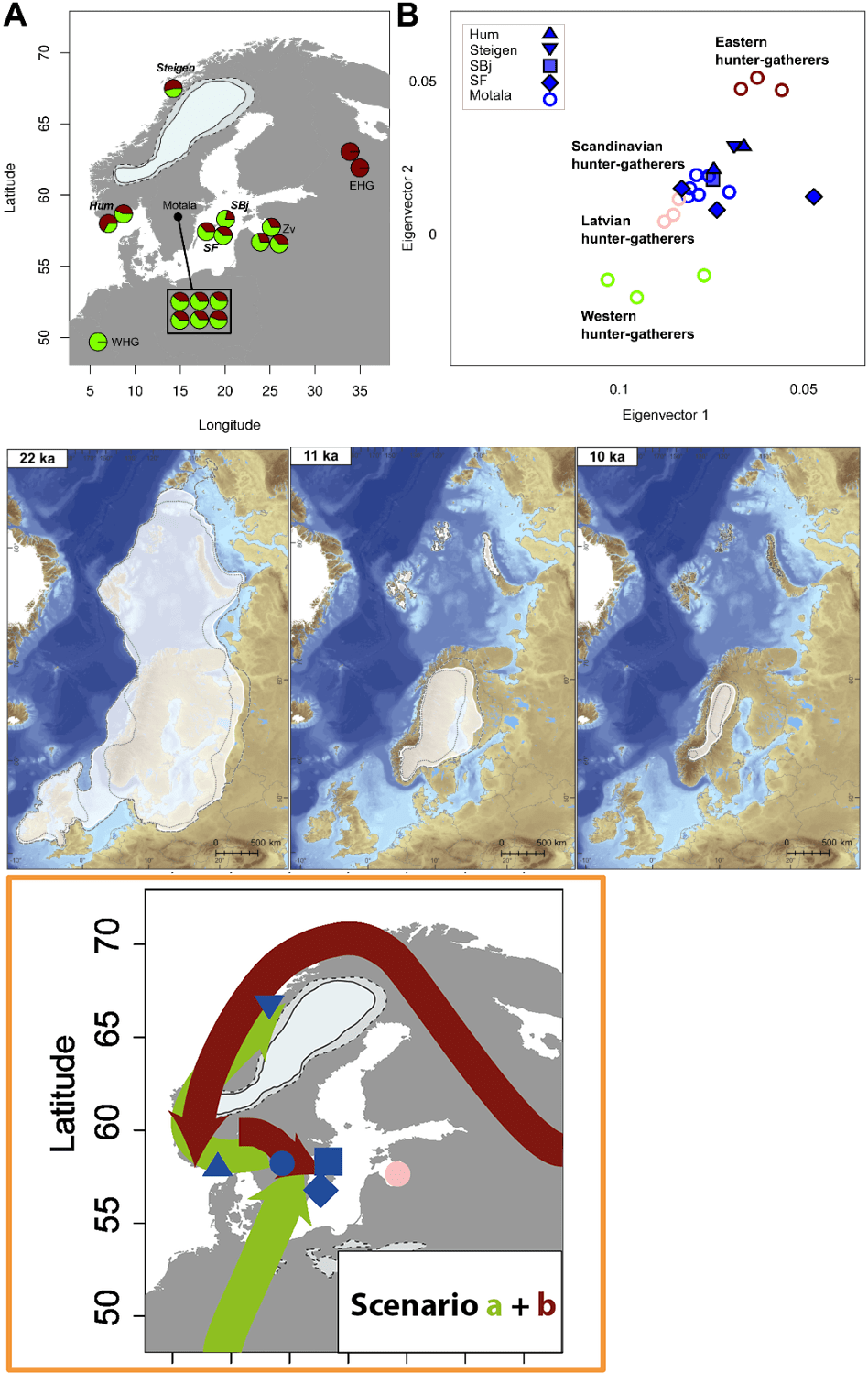
Други рад који се бавио мезолитском древном ДНК јесте рад Хака и сарадника из 2015. године, који су анализирали Y-ДНК 5 мушких скелета из Мотале у Шведској (Haak, et al., 2015). Код свих је установљена I2 хаплогрупа. Узорак I0012, датиран између 5898-5531 п.н.е, припада I2c-L596 хаплогрупи; I0013, датиран између 5898-5531 п.н.е, припада хаплогрупи I2a>M423; I0015, 5898-5531 п.н.е. је I2a>P37; а I0017 припада I2a>L1498.</>
Из приложених радова закључујемо да су мушкарци са мутацијама које су рођачке у односу на I1-M253 били присутни међу првим становницима Скандинавије с тим да нису били репрезентативније заступљени у удјелу мезолитске популације сјеверних дјелова Европе, који су доминантно били I2 хаплогрупе. Како поуздано знамо да је предак данашње I1 популације живио прије 4600 година на подручју централне Европе, о чему ће бити ријечи у наставку рада, закључујемо да поменути мезолитски мушкарци са раним I1 мутацијама нису оставили потомства међу данашњом популацијом, као ни раније поменути са подручја Пиринејског полуострва, дакле ради се о линијама које су изумрле.
НЕОЛИТ
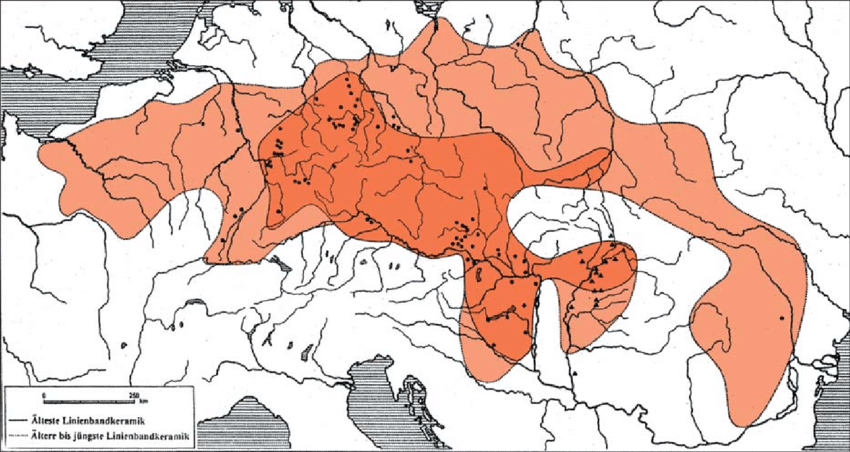
За неолитску револуцију у централној Европи заслужна је Култура линеарне керамике (5500-4500. год. п.н.е.) која је уједно важна и за данашњу I1 популацију. Развила се на подручју Прекодунавске Мађарске гдје је коегзистирала са јужноевропском Старчево-Кереш-Криш културом која је имала развојног утицаја на њу (Banffy, 2000). Захватала је Мађарску, Словачку, Чешку, Пољску, сјеверни Банат, југозападну Украјину, Њемачку, Холандију, Белгију, Француску, Швајцарску и Аустрију. У Прилогу 2 налази се карта на којој је приказана распрострањеност културе. Подручје Културе линеарне керамике генетички су покриле студије Липсона и сарадника као и Сиџињи-Нај и сарадника, Никитина, Матинсона, Гамбе и Хака. Како је у овој култури пронађен најстарији потврђени I1-M253 узорак, за потребе рада предочићемо све анализиране узорке из поменуте неолитске културе ради стицања свеобухватне слике и контекста у коме су живјели I1 мушкарци. Од 38 анализираних мушких скелета само је један припадао I1 хаплогрупи док њих 18 припада G2 (G2a, 18 и G2, 2) хаплогрупи; 6 их је I2 хаплогрупа; 4 су C1a и један је CT хаплогрупа (ближа грана није одређена); два узорка су T1 хаплогрупа; хаплогрупи F припадају 2 узорка (ближа хаплогрупа није одређена), хаплогрупи H 2 узорка, хаплогрупи J2a 2 узорка и један узорак припада R1b хаплогрупи. У Табели 1 (в. чланак у .pdf верзији) приказани су сви досадашњи узорци Културе линеарне керамике.
Из приложеног се уочава да су у Култури линеарне керамике претежно учествовале хаплогрупе анадолских фармера G2a, J2a и T1 док су мање заступљене I2a, I1 и C1a хаплогрупа, својствене првом становништву Европе, ловачко-сакупљачким заједницама које су се прикључиле овој заједници земљорадника. Даље, из ових резултата заједно са раније поменутим мезолитским узорцима, као и из резултата других студија које су се тицале европских неолитских култура, а у којима није пронађена I1 хаплогрупа, извлачимо аналогни закључак да су преци данашњих I1 мушкараца били груписани на ширем подручју Културе линеарне керамике.
Следећа неолитска култура која би била од важности јесте Култура левкастих пехара (4300-2800. год. п.н.е.) која се простирала на подручју данашње Данске, Пољске, Украјине, јужне Скандинавије и у њеном касном периоду до Бјелорусије. Представља прву земљорадничку културу Скандинавије која је коегзистирала са културом ловаца сакупљача. Нажалост она није била у ширем фокусу истраживача који су радили анализе древне ДНК, те је за сада анализирано свега неколико мушких скелета припадника КЛП-а (Култура левкастих пехара), њих 6. Један припада IJK хаплогрупи (ближа хаплогрупа му није одређена), а евидентиран је под шифром I0 802, датиран између 3400-3025. год. п.н.е.; узорак ros5 је I2-пра-S2742, датиран између 3090-2920. год. п.н.е.; I0 551 припада G2a-PF3147 хаплогрупи, 3400-3025. год. п.н.е.; I6677 је H2 хаплогрупа, 3800-3400. год. п.н.е.; I0 807 је I2 хаплогрупа, 3970-3710. год. п.н.е.; и I0 559 је R1b-V2219 хаплогрупа, датиран између 3654-3527. год. п.н.е. Иако није пронађена у ових 6 узорака, присуство I1 хаплогрупе не може се искључити из три разлога. Први је тај што је тренутни узорак сувише мали, друго, КЛП се наслања и временски и географски на Културу линеарне керамике и трећи разлог је тај што у генетичким примјесама тестираних припадника Културе левкастих пехара из Шведске постоји доминантна, надполовична генетичка компонента припадника Културе линеарне керамике (погледати Прилог 3).
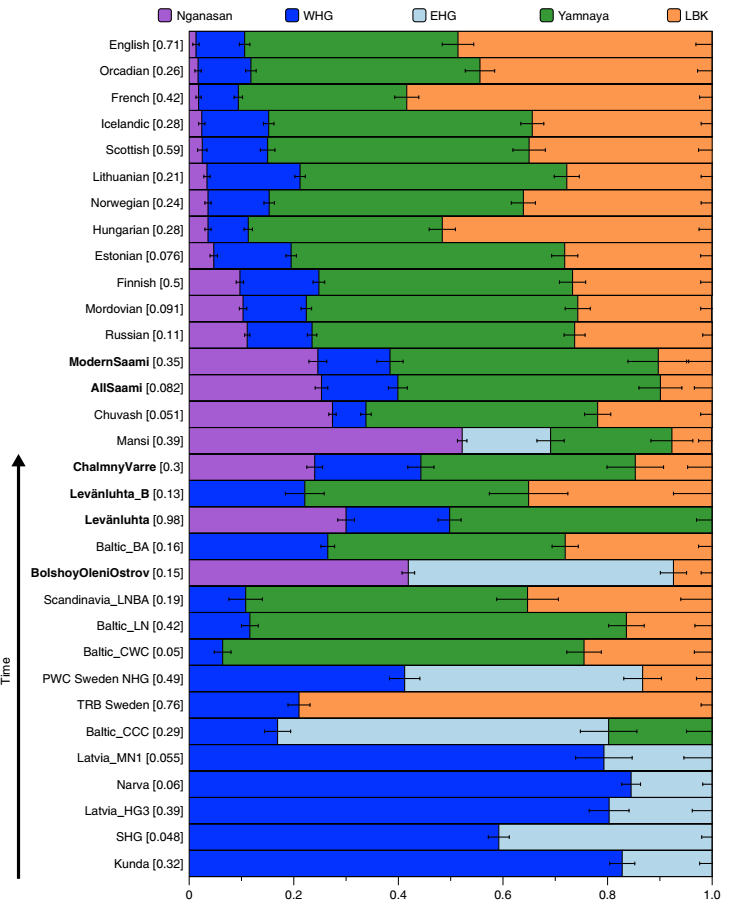
Овде ћемо додати да је у раду Алентофт и сарадника (Allentoft, et al., 2015), пронађен каснонеолитски I1 узорак са југа Шведске заведен под шифром RISE179, датиран у период 2010-1776. год. п.н.е, што додатно потврђује неолитско присуство I1 мушкараца на југу Скандинавије. Узевши приложено у обзир, сасвим сигурно можемо сматрати да су преци данашњих I1 мушкараца насељавали поред централне Европе и јужну Скандинавију, већ током скандинавског неолита, односно од Културе левкастих пехара. Дакле подручје где су били груписани током неолита се налазило на ширем подручју ове двије културе. Како је заједнички предак данашњих I1 мушкараца живио неких 2000 година након Културе линеарне керамике, односно неколико стотина година после Културе левкастих пехара, ми не можемо са сигурношћу тврдити ништа више од горе наведеног. Остаје непознато који догађаји су утицали на страховито уско грло ове хаплогрупе.
ЕНЕОЛИТ
На сјеверу Европе енеолит почиње нешто после 2800. године п.н.е и завршава се око 1700. године п.н.е, када наступа бронзано доба. Култура врпчасте керамике (2900-2350. год. п.н.е.) је карактеристична култура касног неолита и енеолита. Главна је праисторијска индоевропска култура сјеверне и средње Европе, простирала се од Холандије и Швајцарске на западу, па на исток кроз средњу Европу до горњег тока Волге и средњег тока Дњепра, те на сјевер у Скандинавију.
Погодна клима, налик оној данашњој са сјевера Француске, у Скандинавији је трајала од 2700-850/650. год. п.н.е, што је допринјело насељавању припадника Културе врпчасте керамике (Kane, 2019).
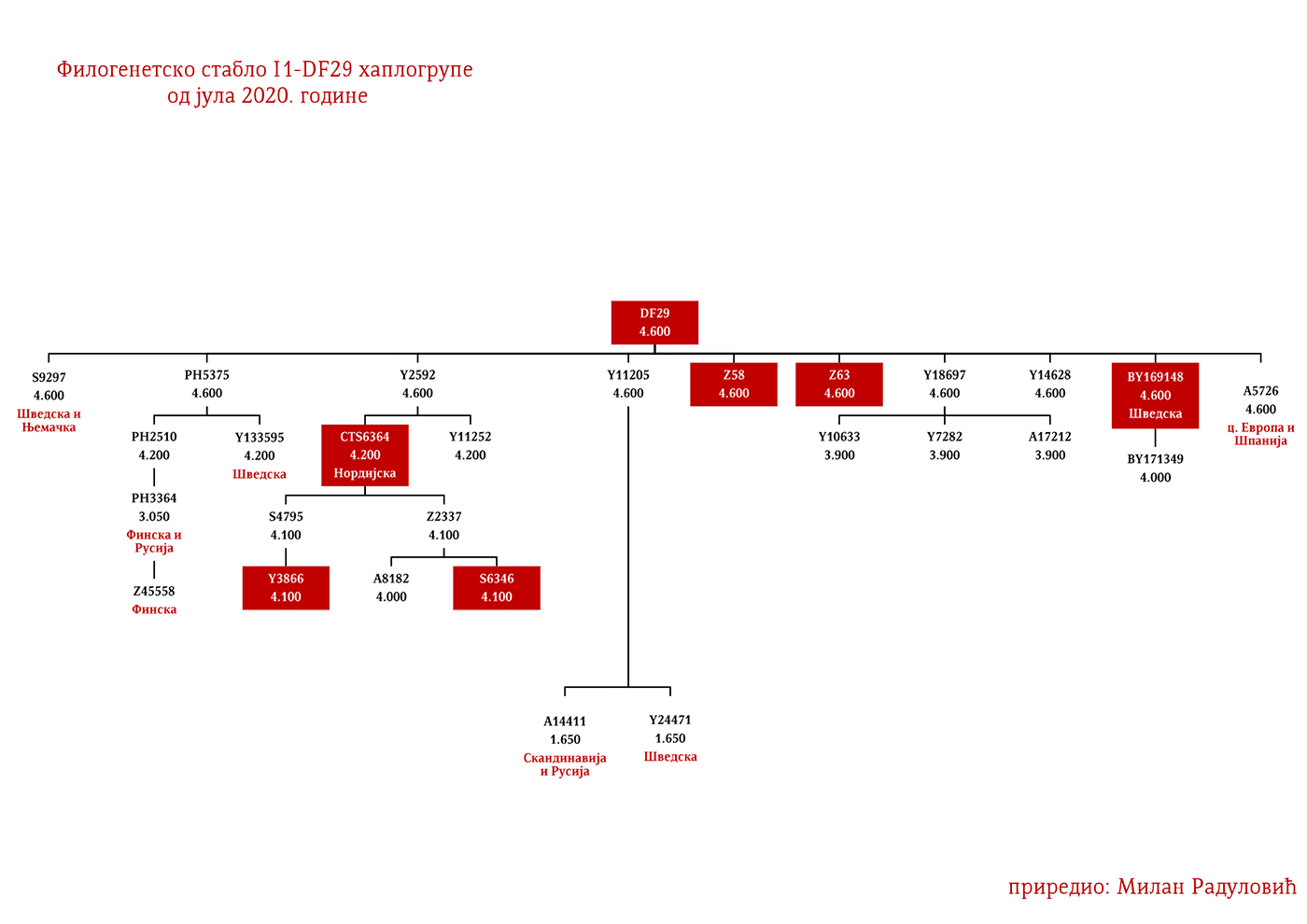
Да бисмо стекли цјеловиту слику прикупили смо све археогенетичке радове који су се бавили овом културом и анализирали Y-ДНК њених припадника. У Табели 2 (в. чланак у .pdf верзији) налази се комплетан преглед свих 37 прикупљених узорака. У овој култури су претежно били заступљени R1a и I2a мушкарци, а мање су заступљени припадници хаплогрупа R1b и G. Мушки I1 скелет са локалиетеа Елсе (Ölljsö) налази се под шифром oll009 датиран између 1930-1750. год. п.н.е, а према анализи нашег стручњака за древну ДНК Милана Рајевца, која је у форми детаљне анализе урађена наменски за потребе овог чланка, oll009 припада I1a-DF29 хаплогрупи (Рајевац, 2020). Други интересантан узорак је већ поменути RISE179 са локалитета Абекос (Abekås I), који је такође I1, са подручја Културe врпчасте керамике.
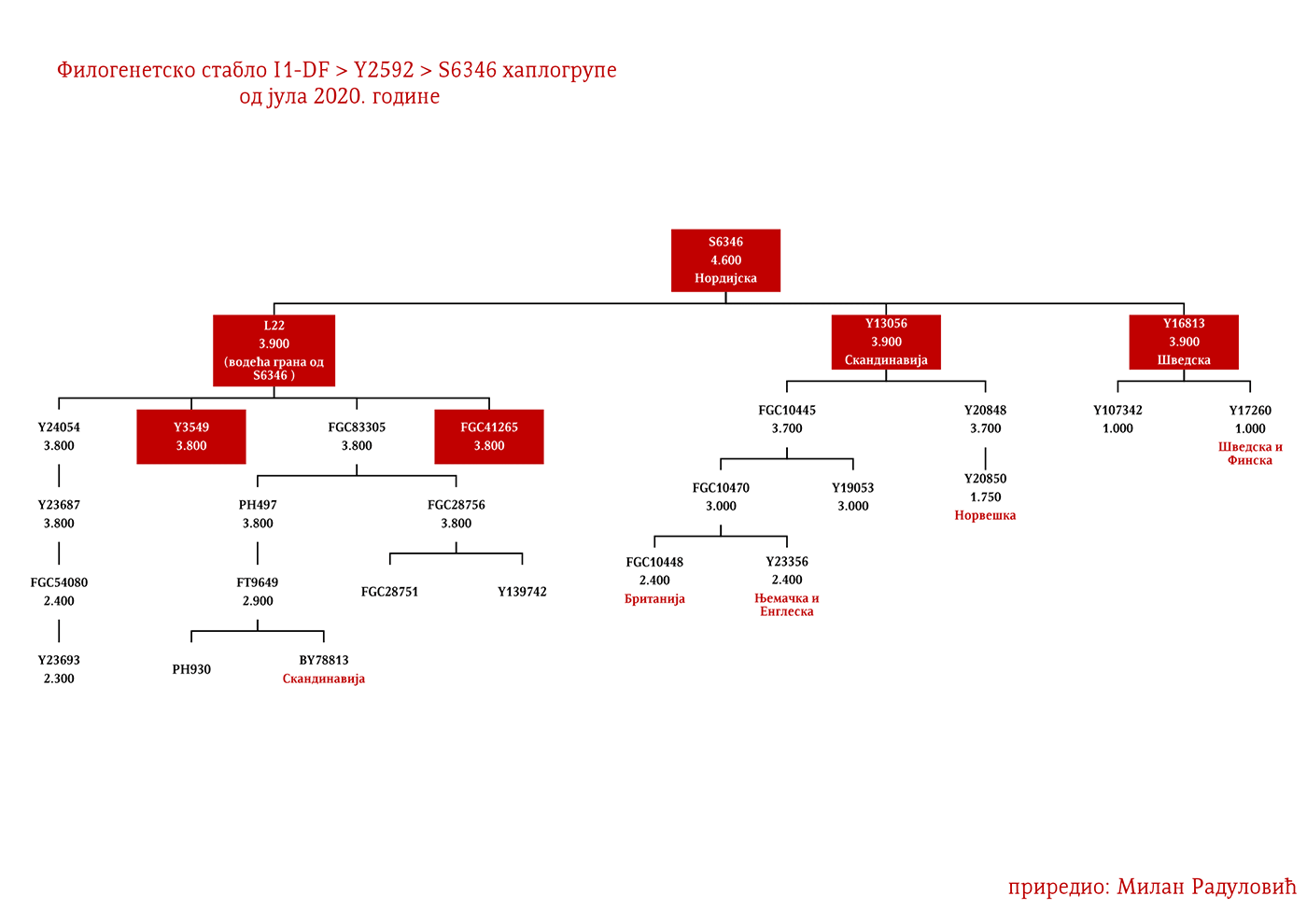
Дакле, од 37 узорака њих четири је из Шведске, а од тога два узорка припадају I1 хаплогрупи, међутим они нису били део Културе врпчасте керамике већ коегзистирајуће каснонеолитске културе. Имајући у виду касније налазе, нема сумње да су се утопили у ову културу и да су чинили добар дио њеног огранка познатог под називом Култура бојних сјекира. Са доласком припадника Културе врпчасте керамике у Скандинавију, наступио је енеолитски период без неке посебне неолитске транзиције и најављено је бронзано доба. Уједно вриједи поменути да је ово вријеме почетка ширења I1 популације, односно настанка грана I1-CTS12768 (која за сада није уочена од стране ISOGG), I1-Z17954 и I1-DF29 која се убрзо разгранала на низ огранака. Са подручја сјевера централне Европе следеће гране од I1-DF29>Z63 су започеле ширење централном и западном Европом: I1-Z63>BY62 и BY151>FGC81364, као и гране од I1-Z58, I1-Z58>A6867 и I1-Z58>PH4482, док I1-L22 започиње нагло ширење међу протонордијском популацијом, а остале гране I1 доживјеће ширу експанзију централном Европом махом током периода 1800-1200. године п.н.е, судећи по времену настанка грана, њиховој разноврсности и заступљености, те времену до заједничког претка.
Већ у наредном периоду после 1700. године п.н.е. уследиће нови талас Индоевропљана који ће Скандинавији донијети германски карактер.
БРОНЗАНО ДОБА
Нордијско бронзано доба почиње 1700. године п.н.е. и завршава се 500. године п.н.е, а њега карактерише нови талас досељеника из централне Европе оличених у хаплогрупама карактеристичнијим за германске народе. Тада започиње нагло ширење I1 хаплогрупе на тлу централне Европе, са интензивирањем комуникације са континенталним дијелом. Бродови постају главна средства комуникације и трговине, њихов значај види се поред осталог и у томе што су Нордијци током бронзаног доба правили споменике у виду „камених бродова“ (Kane, 2019) на гробним хумкама.
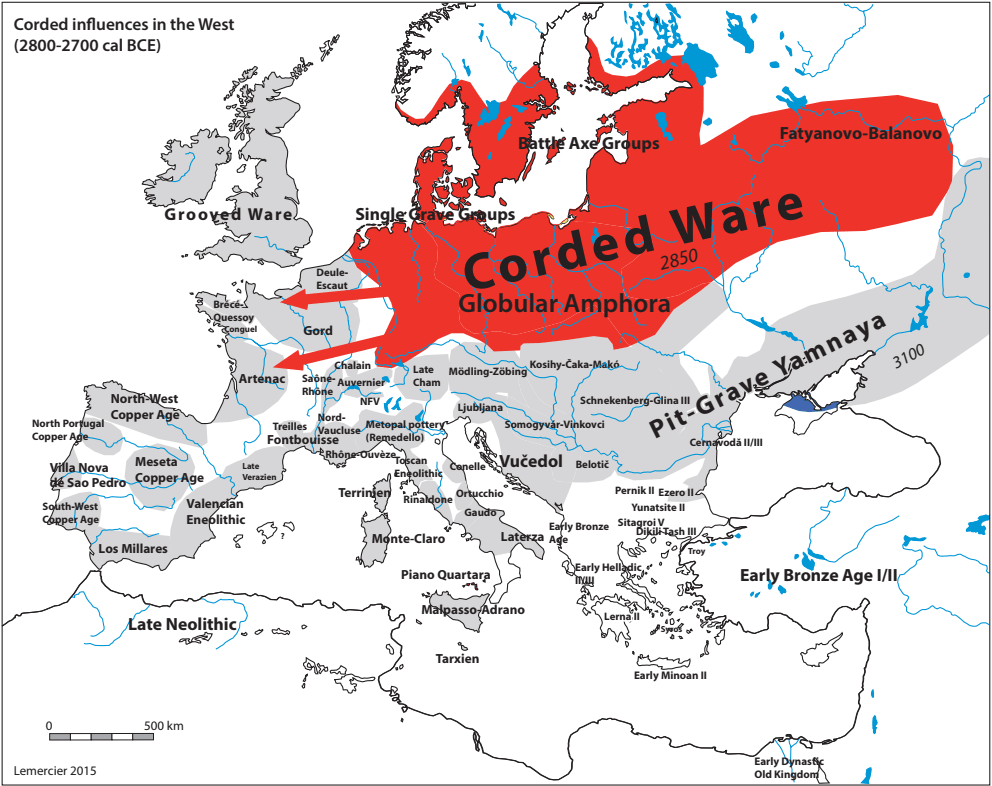
Други талас Индоевропљана врло је вјероватно последица експанзије Уњетичке културе (2300-1600. год. п.н.е.) са простора данашње Њемачке. Одатле су се припадници R1b-U106 хаплогрупе проширили на сјевер, гдје су први датирани у 1400. год. п.н.е. према резултатима из студије Алентофт и сарадника. Сама Уњетичка култура слабије је покривена генетичким студијама, тестирано је свега 14 (I5043, I0804, I7197, I0114, I0116, I7199, I4884, I5037, RISE145.SG, I5042, RISE431.SG, I7203, I7196, I7202, узорци из поменутих студија за Уњетичку културу) њених мушких припадника. Углавном су је судећи по узорку (Olalde, et al., 2018; Allentoft, et al., 2015; Mathieson, et al., 2015) сачињавали I2a2, R1a и R1b мушкарци, са мањим удјелом G2a хаплогрупе. Током поменутог периода одиграла се прва етногенеза германских народа и у њој су учествовали припадници I1 хаплогрупе. Уједно ово је, како смо претходно навели, период наглог ширења њених подграна. У овом периоду су следеће подгране извршиле уочљивије позиционирање: Z63 која је махом концентрисана у централној Европи на простору данашње Њемачке, и изостаје у нордијској популацији; Z58 која је концентрисана у западним дјеловима централне Европе; L22 која је груписана у Скандинавији, као и Z2336, док је M227 смјештена источније. Како се вријеме ширења подудара са етногенезом германских народа, вриједи погледати карту из Прилога 5 на којој је приказана експанзија Германа, која је релевантна за I1 хаплогрупу.
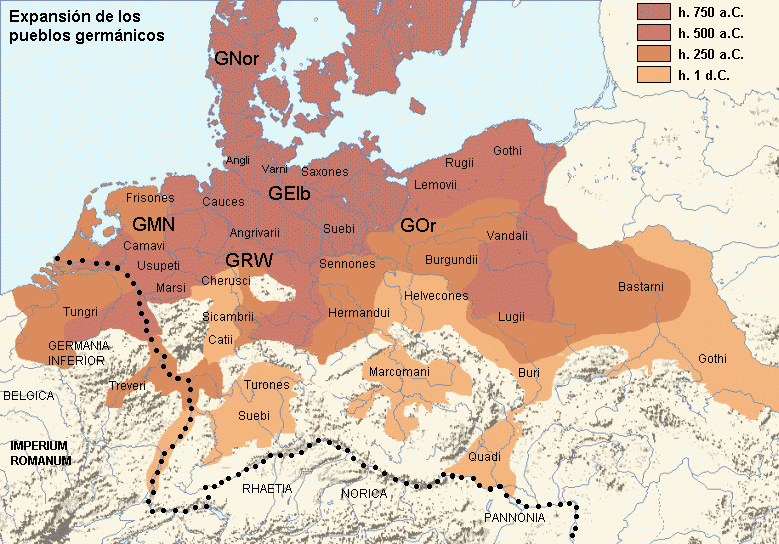
Од 6 узорака древне ДНК из нордијског бронзаног доба из Шведске, 4 су I1 хаплогрупе и то су узорци RISE179, RISE207, RISE210, RISE175, док су 2 R1b хаплогрупе. На миграције из правца југа Скандинавије ка централној Европи вјерује се да је битно утицала нагла климатска промјена која се десила у периоду између 850. и 650. године прије нове ере, када је наступило нагло захлађење (Kane, 2019).
ГВОЗДЕНО ДОБА И АНТИКА
Како је фокус хаплогрупе на Скандинавији, гвоздено доба дијели се на предримско гвоздено доба које почиње 500. године п.н.е. и траје до 1. године нове ере, други период је римско гвоздено доба које траје од 1. до 400. године н.е, затим следи германско гвоздено доба од 400. до 800. године.
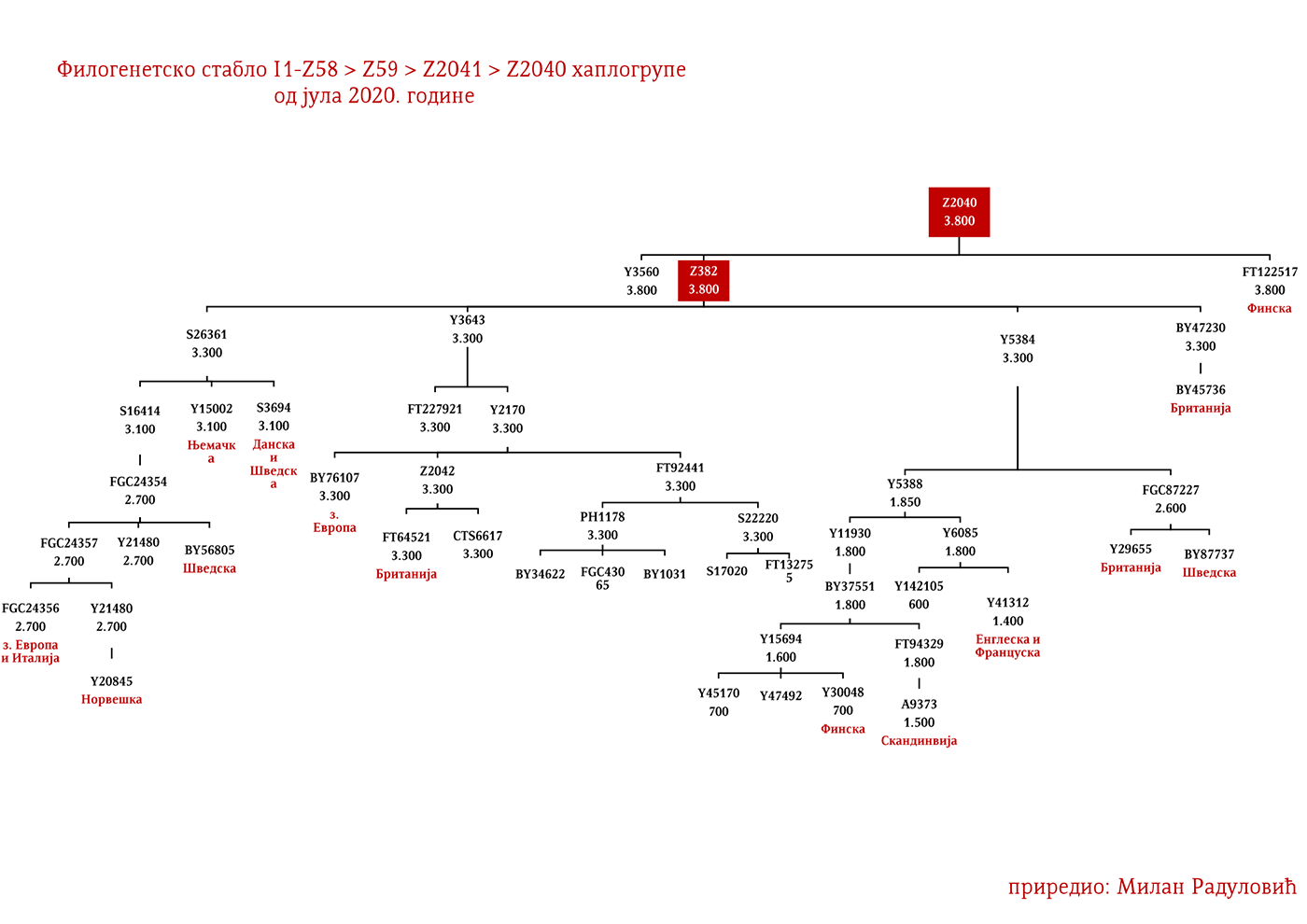
Најстарији узорак којим располажемо из гвозденог доба је са југа Француске из Бесана, а припадао је латенској култури (450-1. год. п.н.е.). Налази се под шифром BES1248 и датиран је између 200-50. год. п.н.е. (Brunel, et al., 2020), затим VK521 из Данске (Margaryan A. , et al., 2019), припадник I1-Z58>Z2040 хаплогрупе који је датиран у 300. годину. У централној Европи из овог периода евидентирани су узорци AED_249, STR_241 и STR_486 из рада Верамаха и сарадника (Veeramah, et al., 2018), који потичу из Баварске и датирани су у период између 460-530. године. Код ових узорака карактеристично је то што су сви имали потпуне сјеверноцентралне европске генетичке примјесе (Fig. 3. из студије). Узорак AED_249 припадао је подграни I1-DF29 (xP109), STR_241 је I1-DF29 (xM227, Z138, Z60) док је STR_486 I1-M253 (xP109). Затим имамо узорак KO_55 из Пољске који припада вилбарској култури (Zenczak, et al., 2017), датиран у 100-300. годину, који припада подграни I1-Z63>L1237 (Рајевац, 2020). У Мађарској на локалитету Солад (Szólád), на узорку од 21 скелета један је I1-P109>FGC21810 (шифра узорка је SZ45), док су остали доминантно R1b и I2a2 (Amorim, et al., 2018) мушкарци. На простору Мађарске нађен је још један I1-M253 узорак али из раног аварског периода, SzO/540 (Neparáczki, et al., 2019). Приложени узорци из радова доводе до закључка о разноврсности I1 подграна у централној Европи која одговара њиховој данашњој распрострањености, што се подудара са раније написаним закључцима у вези ове хаплогрупе. Узорци Лангобарда из касноантичког периода из Италије, R110 који припада подграни I1-Z63>Y7234 и CL63 који припада подграни Y2245, заједно са BES1248 са југа Француске, свједоче о миграцијама германских племена. Англосаксонски узорак NO3423 са подручја Велике Британије, који припада подграни I1a-DF29 (xZ63, M227,Y3603, Z2041, Z138, CTS7362, Z141), с друге стране свједочи о западним досезањима I1 мушкараца током касноантичког периода.
SNP МУТАЦИЈЕ
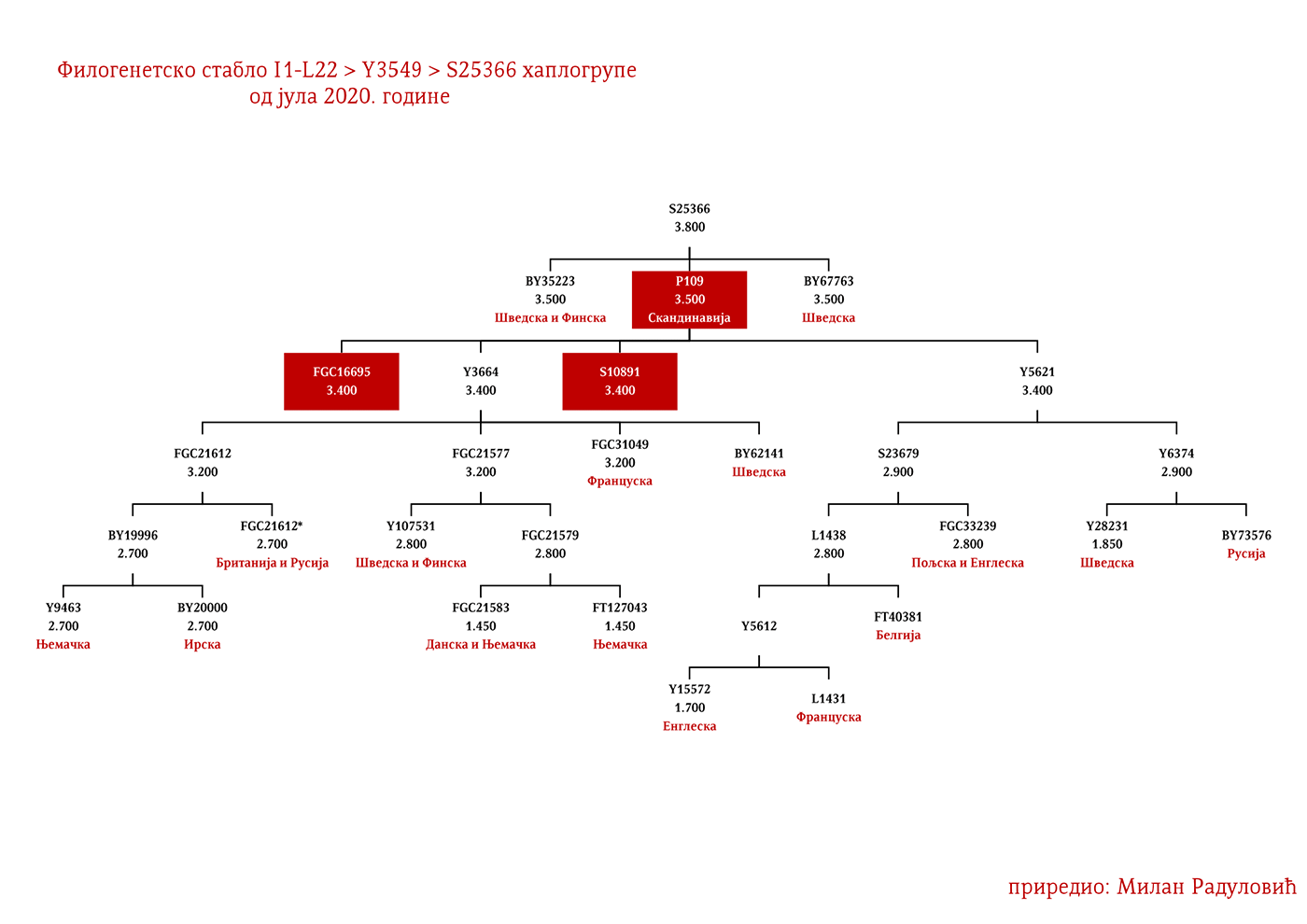
Хаплогрупу I1 дефинишу снипови M253, P30, P40, M307 и M450 (Karafet, et al., 2008). Она се према тренутном ISOGG стаблу 2019 дијели на три гране: I1a коју дефинише снип DF29, I1b дефинисана Z131 и I1c дефинисана Z17943 снипом (ISOGG, 2020). Овде ћемо констатовати да ISOGG касни са освјежавањем стабла, јер Z17943 и Z131 имају заједничког претка Z17954, а који је на истом филогенетском нивоу са DF29 и новоткривеном линијом CTS12768, стога би у будућој освјеженој верзији стабла Z17954 требала носити ознаку I1b док би CTS12768 носио ознаку I1c.
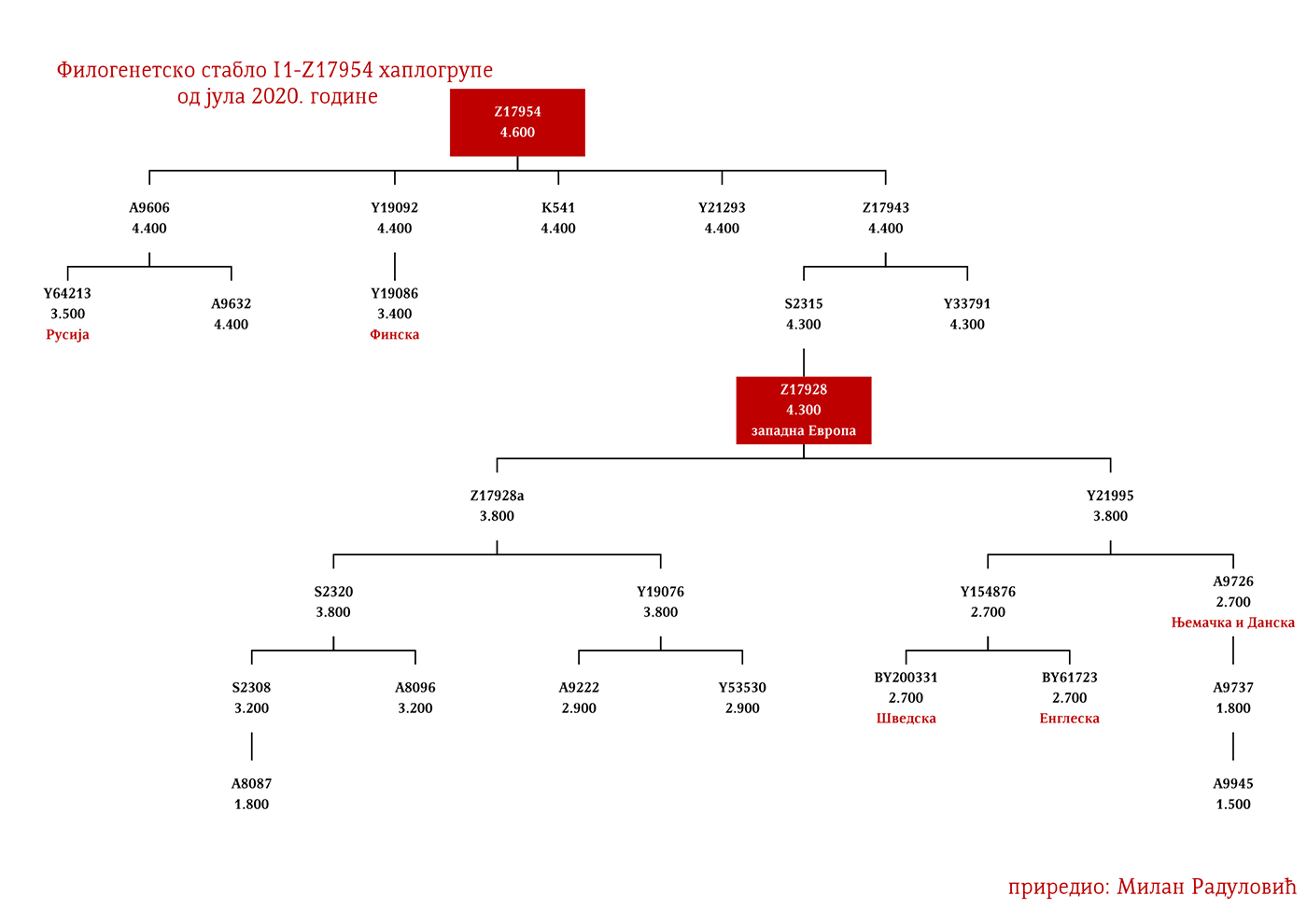
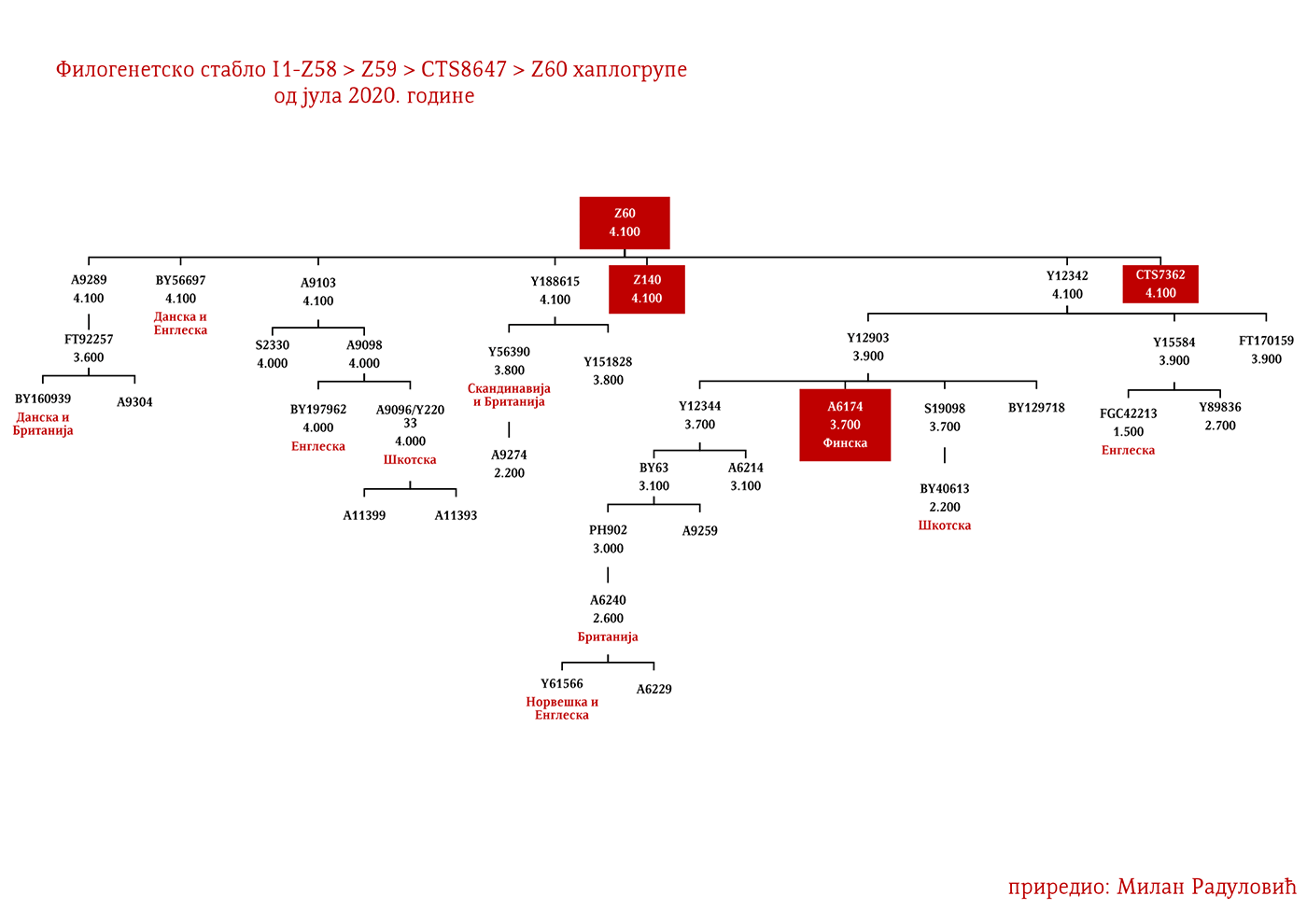
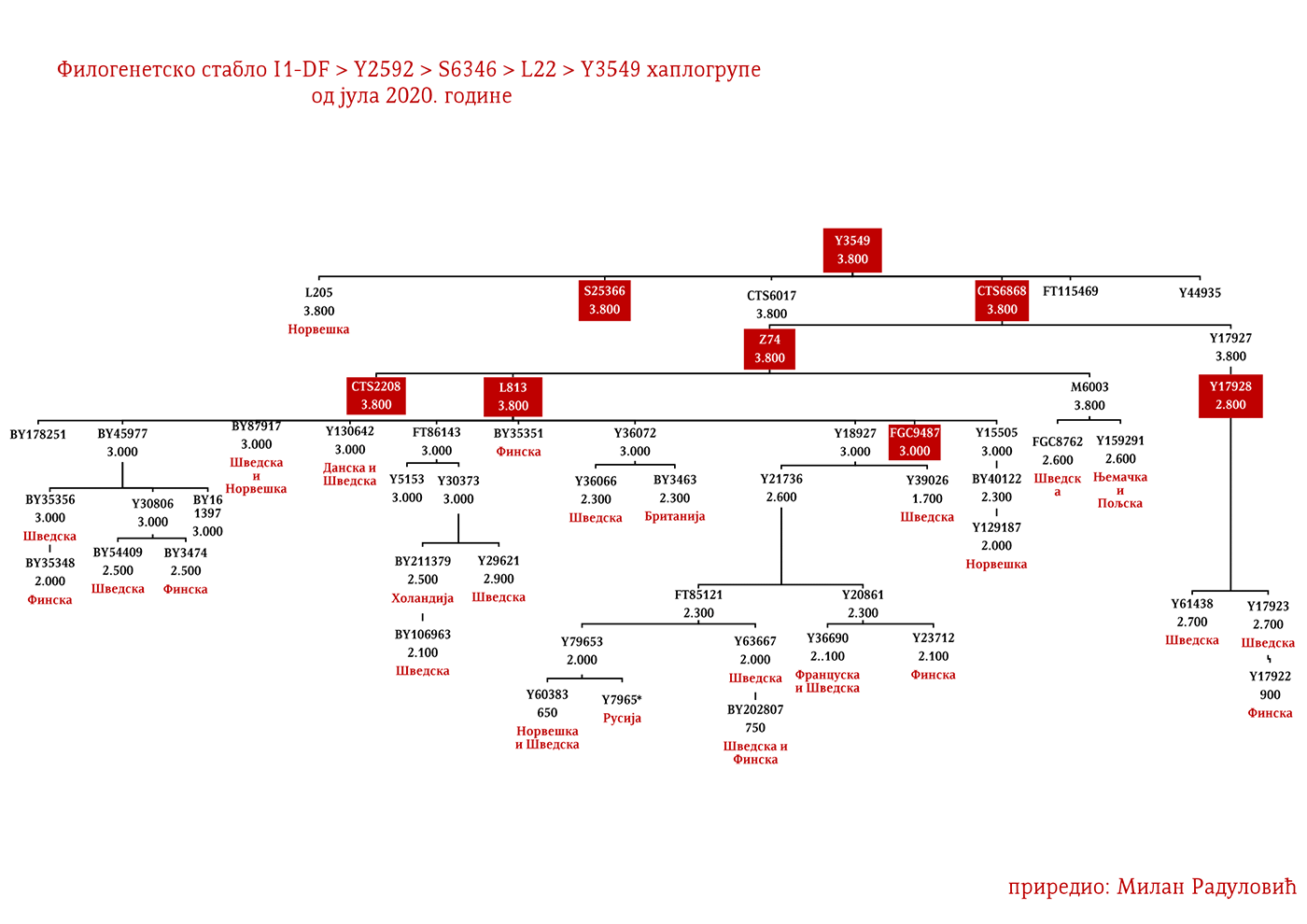
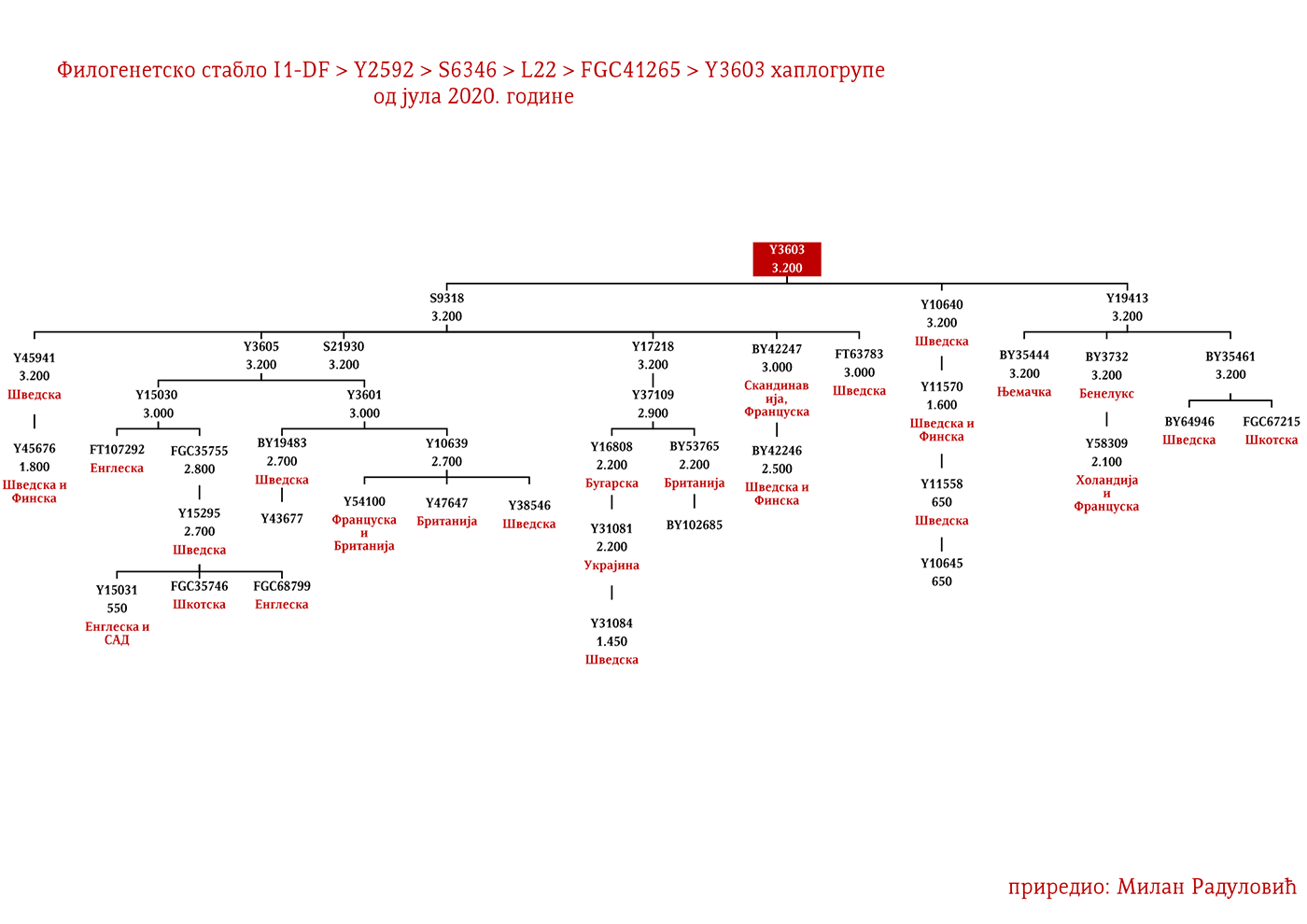
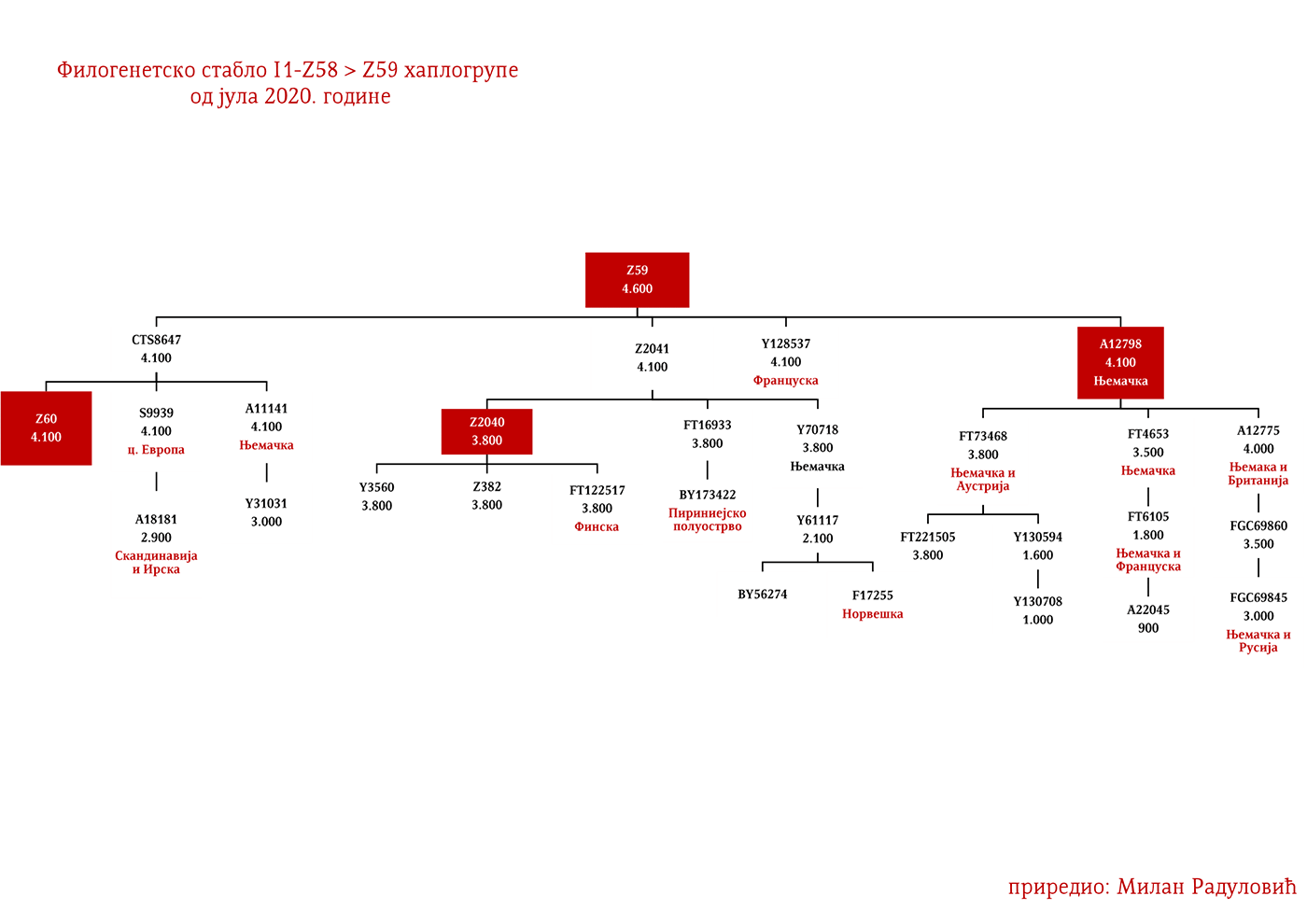
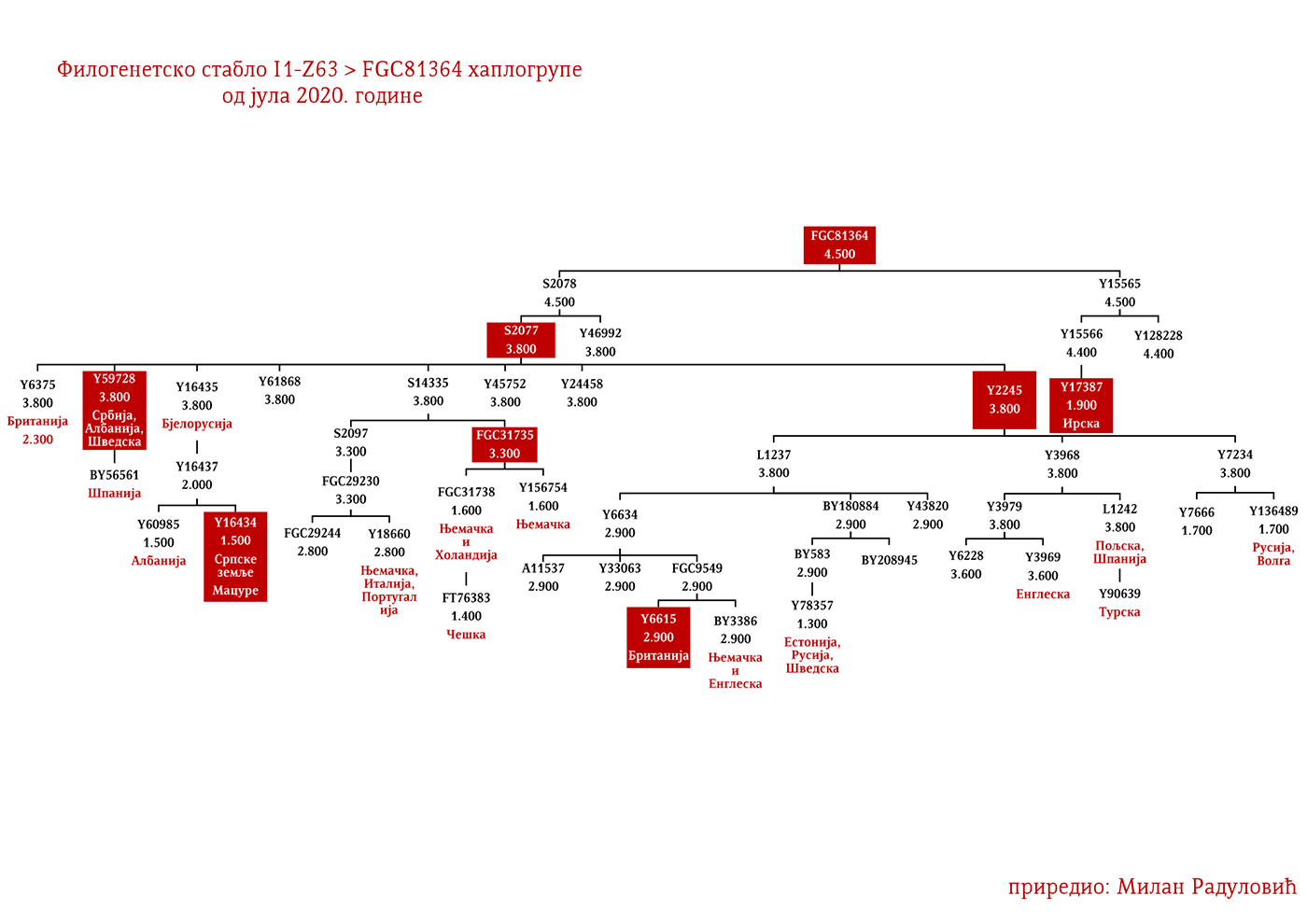
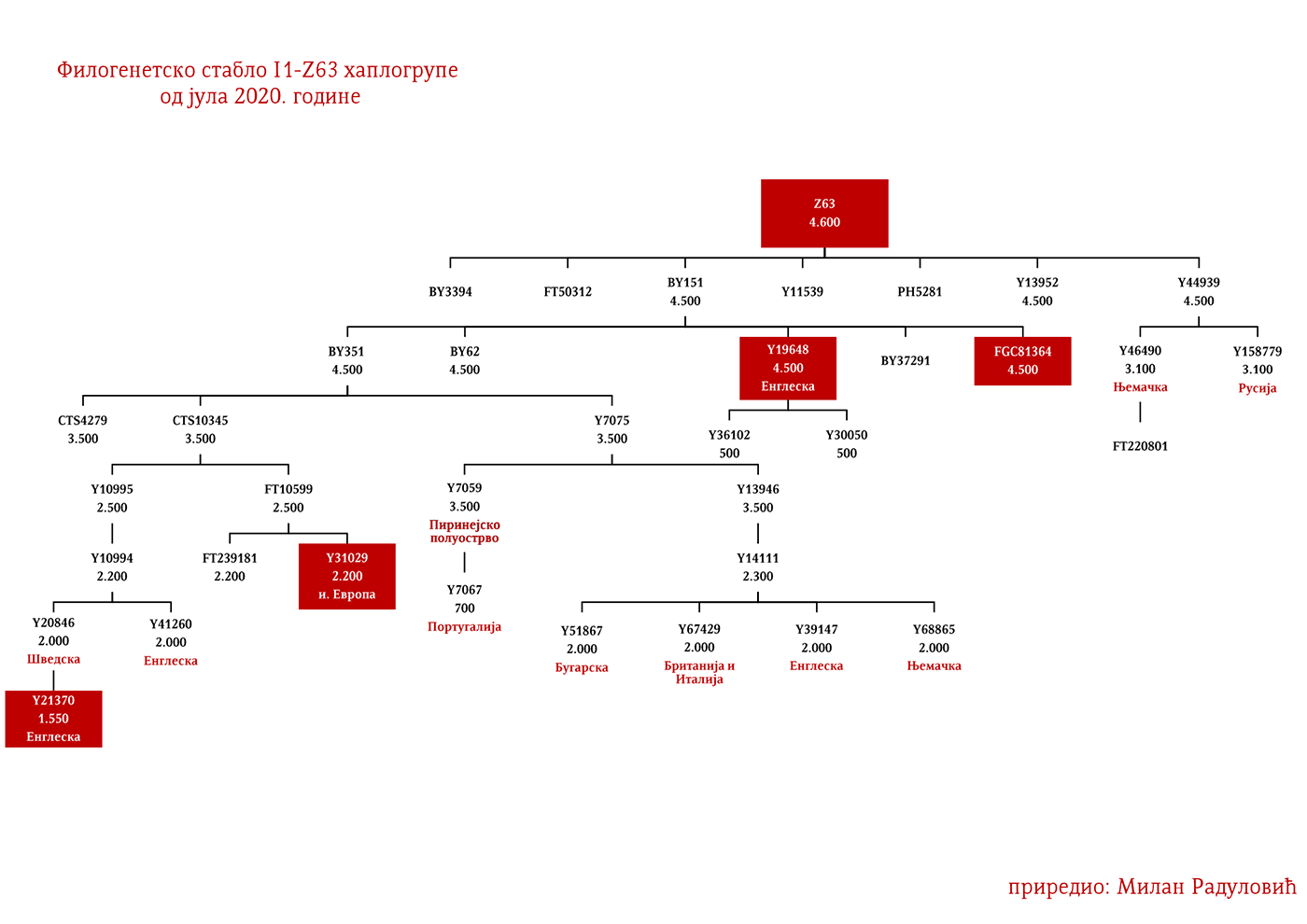
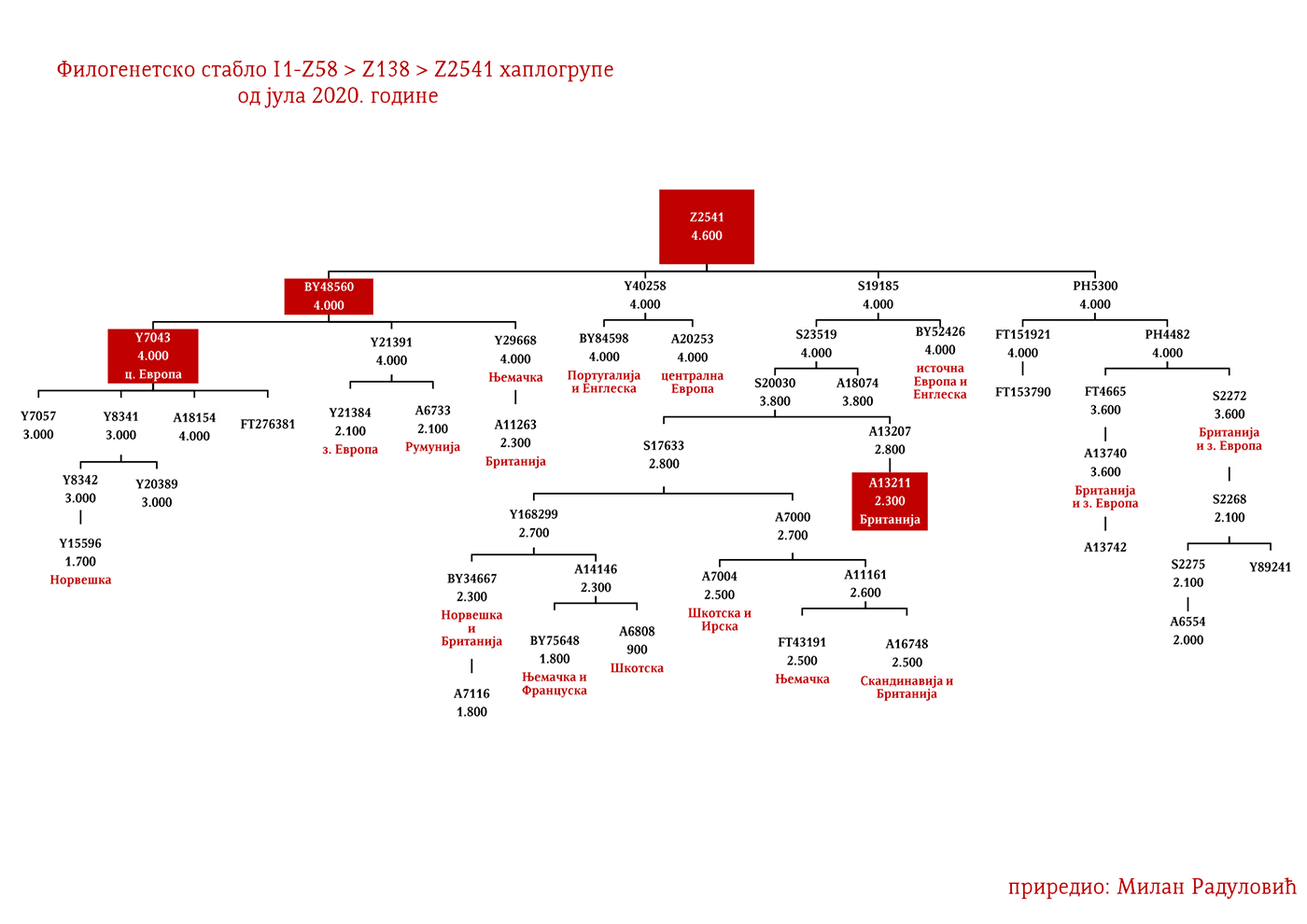
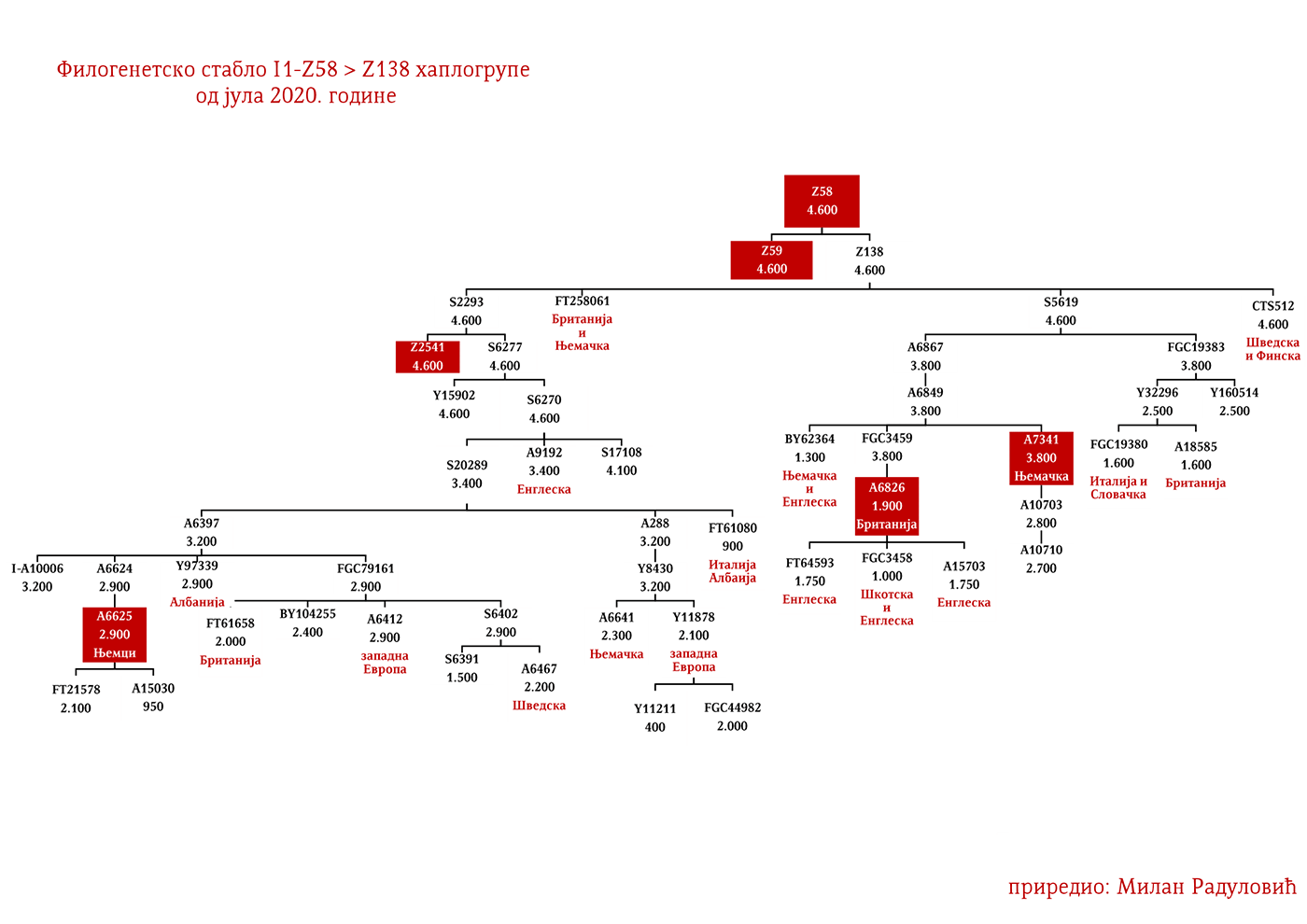
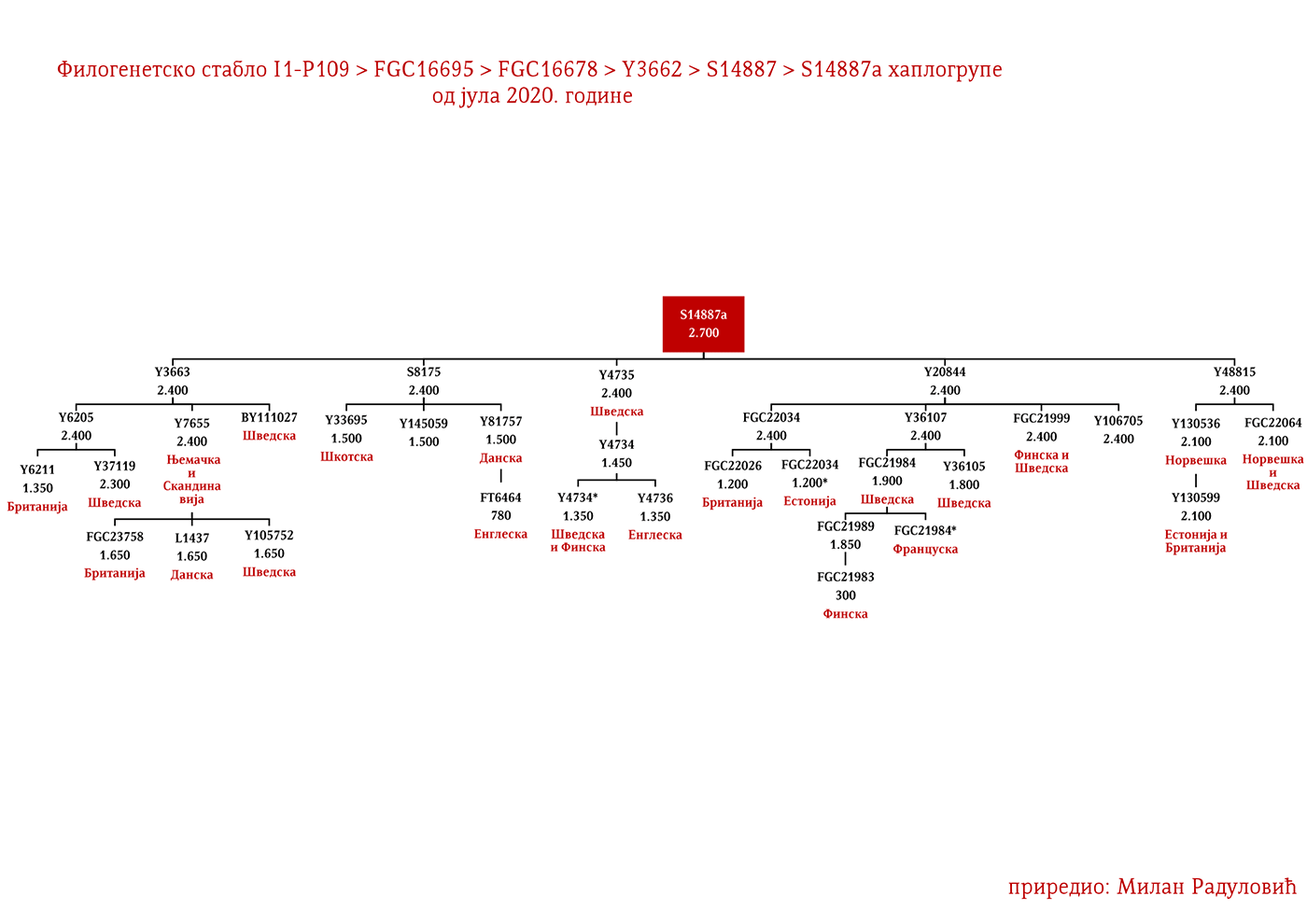
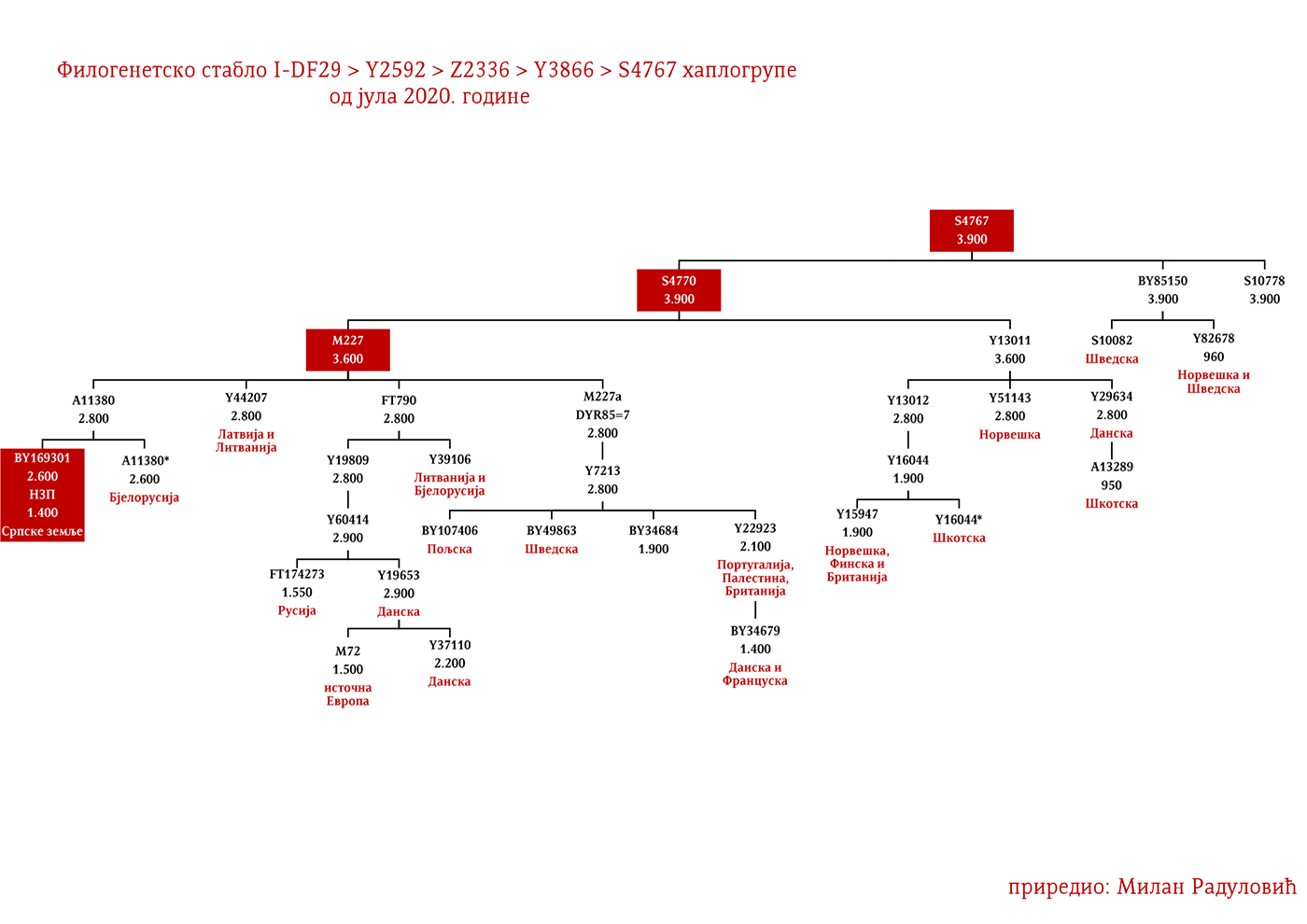
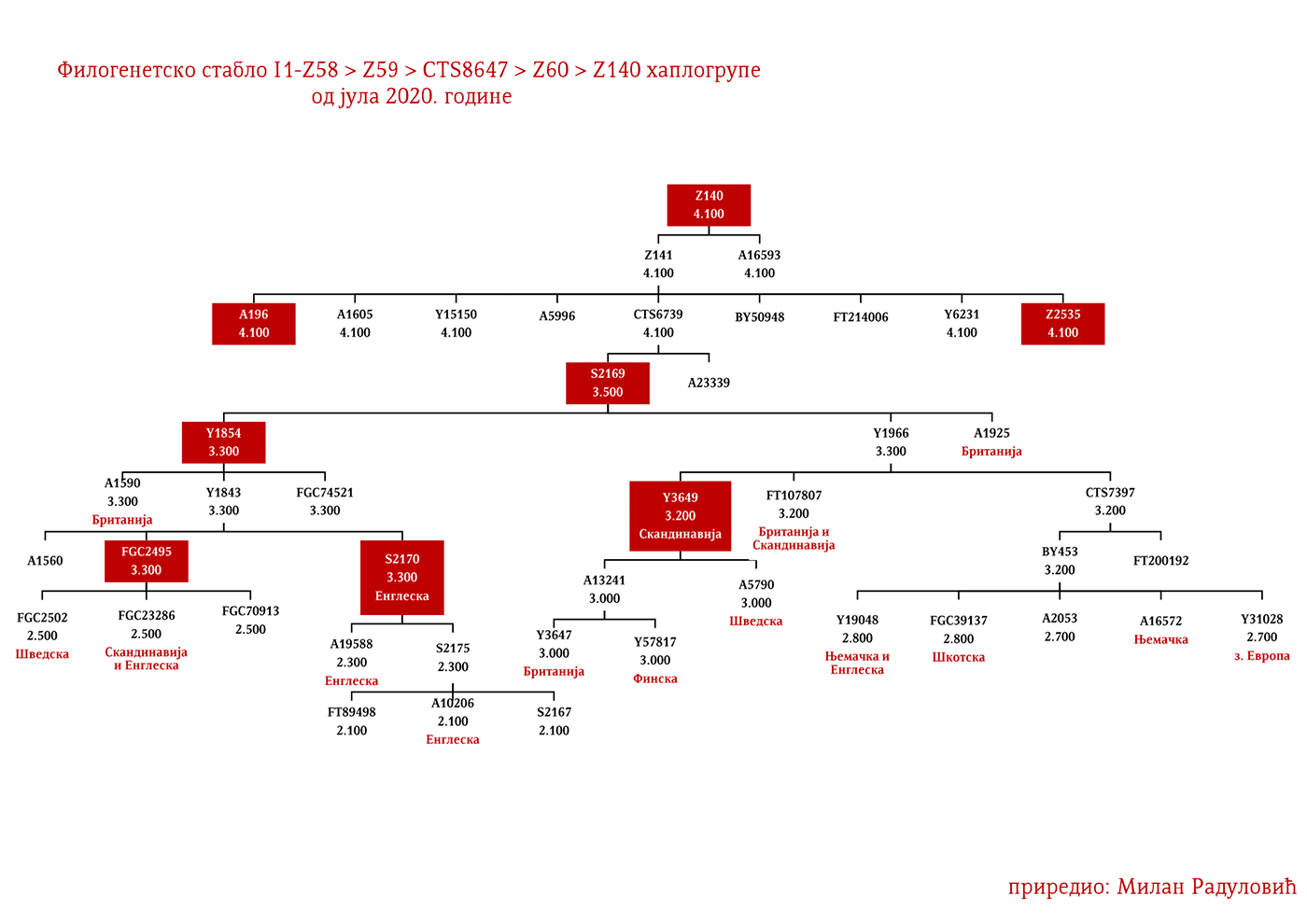
ЗАСТУПЉЕНОСТ И ПОДГРУПЕ
Хаплогрупа I1 заступљена је у Европи гдје на сјеверу континента биљежи највећу заступљеност и разноврсност грана хаплогрупе. Процентуално гледано најзаступљенија је у нордијским и германским земљама. У Норвешкој је заступљена са 39%, Шведској са 35%, Исланду 34%, Данској 32%, Финској 23%, Њемачкој 17%, Холандији 16%, Аустрији, Белгији, Естонији 12%, Великој Британији 11%, док је код српске популације заступљена са 8% (према подацима Српског ДНК пројекта).
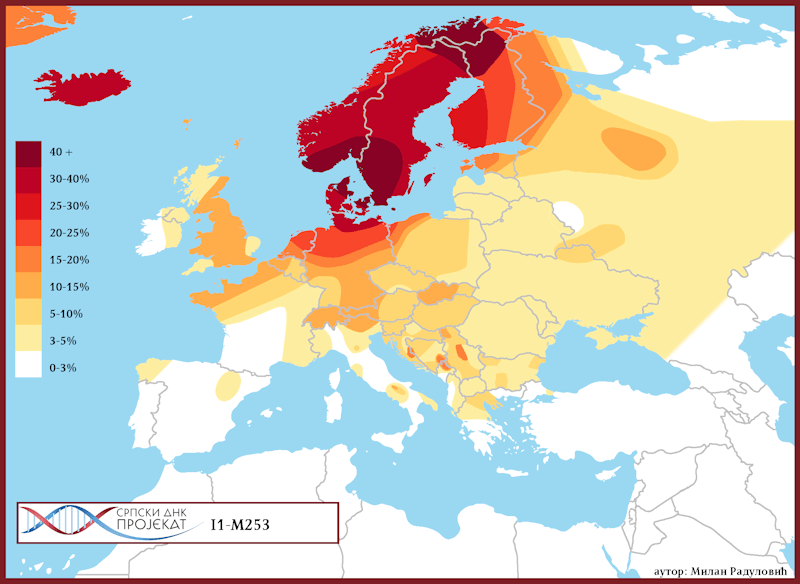
Детаљан преглед заступљености хаплогрупе I1 у европским популацијама можемо видети у Прилогу 7 испод.
У .pdf верзији дат је детаљан приказ грана хаплогрупе, као и њихово географско и етничко гравитирање, при чему још једном скрећемо пажњу да је већина подграна заступљена и нордијским земаљама и код германских народа.
СТАРИ НАРОДИ
Хаплогрупу I1 можемо повезати са германским народима, она је учествовала у њиховој етногенези заједно са R1b-U106, R1b-L238, I2a2-M223, R1a-Z284 и R1a-L664 (хYP942) хаплогрупом, што је својевремено и потврђено археогенетиком.
ВИКИНЗИ
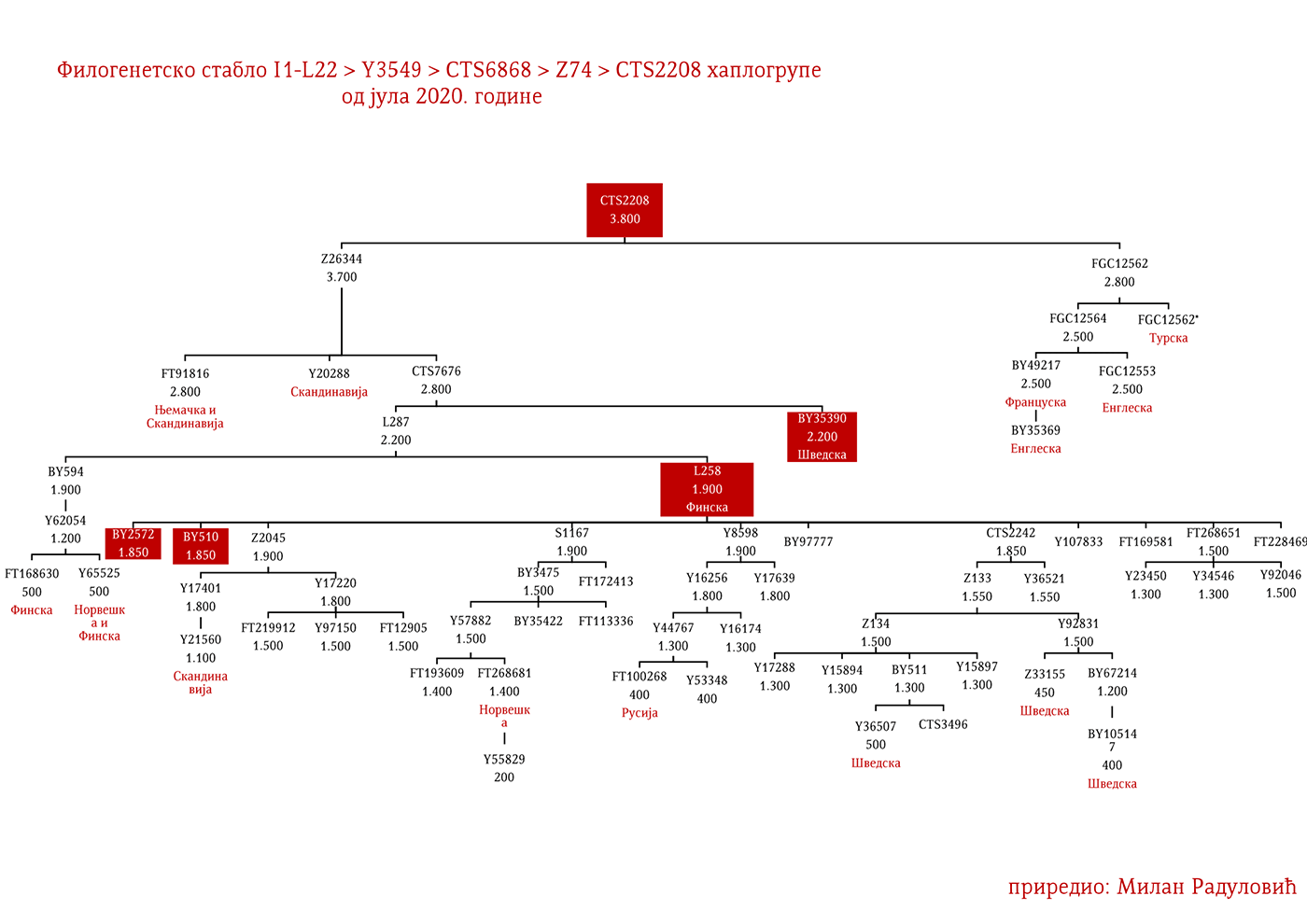
Представљају групу народа са сјевера Европе из земаља Данске, Норвешке и Шведске који су најприје започели ратничке походе ка западној Европи, а нешто касније ка истоку према словенском свијету (гдје се традиционално називају Варјазима). Викиншко доба, како то називају западноевропски историчари, почиње 793. године са нападом Викинга на манастир Линдисфарн на сјеверу Енглеске, а траје до 1066. када су Нормани у битци код Хејстингса поразили енглеског краља Харолда Годвинсона и постали владајућа елита. У недавно објављеном раду о Викинзима, на узорку од 296 мушких скелета (Margaryan A. , et al., 2020), 92 су припадала I1 хаплогрупи, што је трећина од укупног узорка. Од тог броја (92), 37 скелета, односно 40%, припадало је I1-L22 линијама, чиме је потврђено писање ранијих истраживача, Јерковића (Јерковић, Y-ДНК хаплогрупа I – најчешћа код Срба, 2012), Нордветa (подјела на „Norse” и „Ultra-Norse” хаплотипове L22+) и Хеја (Hay, Origins and history of Haplogroup I1 (Y-DNA), 2016) да је L22 најтипичнија викиншка хаплогрупа. Поред L22+ линија, друга по учесталости је Z59+ линија која би иначе претежно била добар кандидат за западногерманске народе, Aнглосаксонце. Упадљиво је одсуство Z63 линија, што и не чуди с обзиром да имају готово искључиво континентални карактер. Пронађена су и два M227 узорка, очекивано на истоку Шведске сходно основаној претпоставци да су углавном учествовали у готским миграцијама.
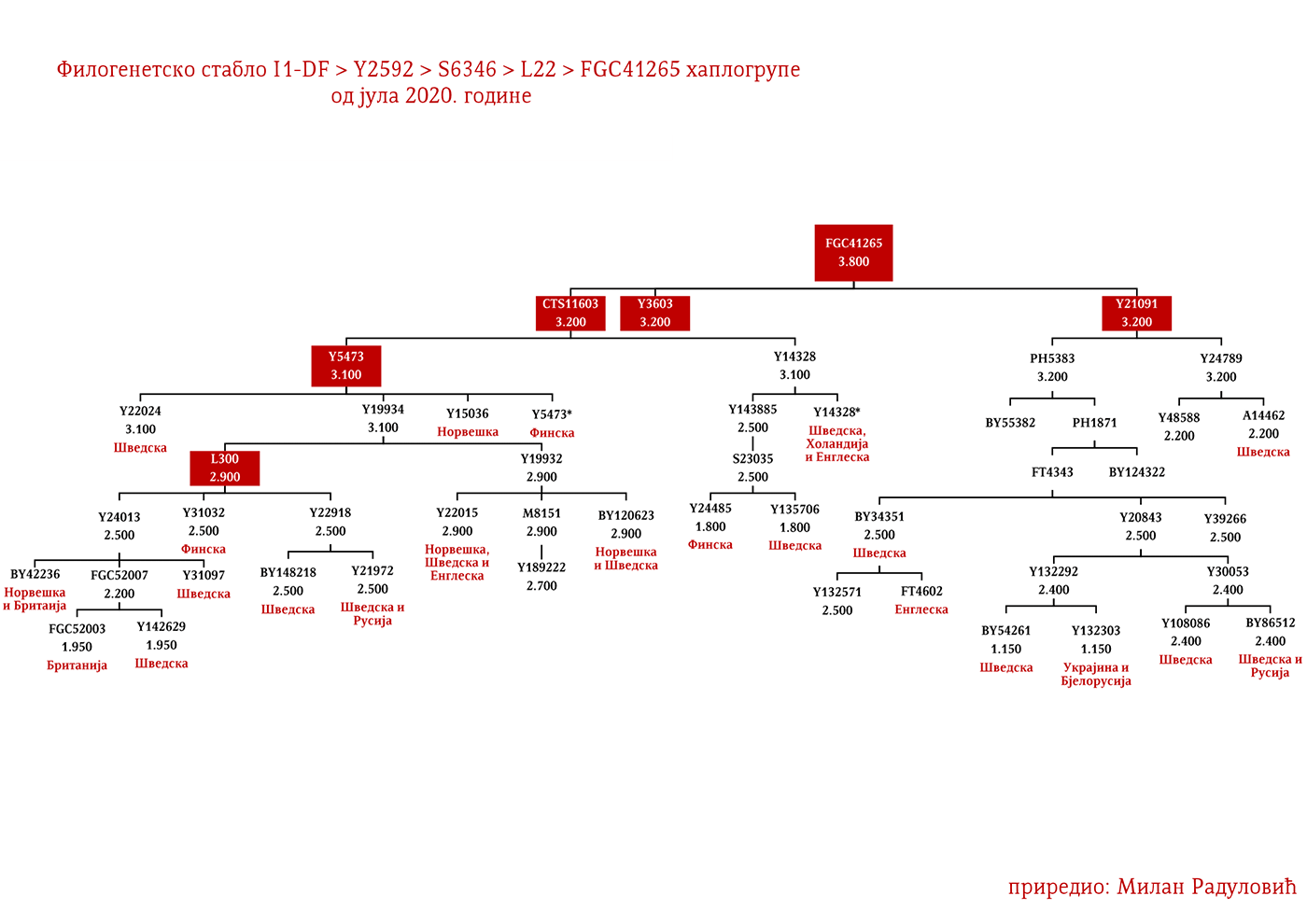
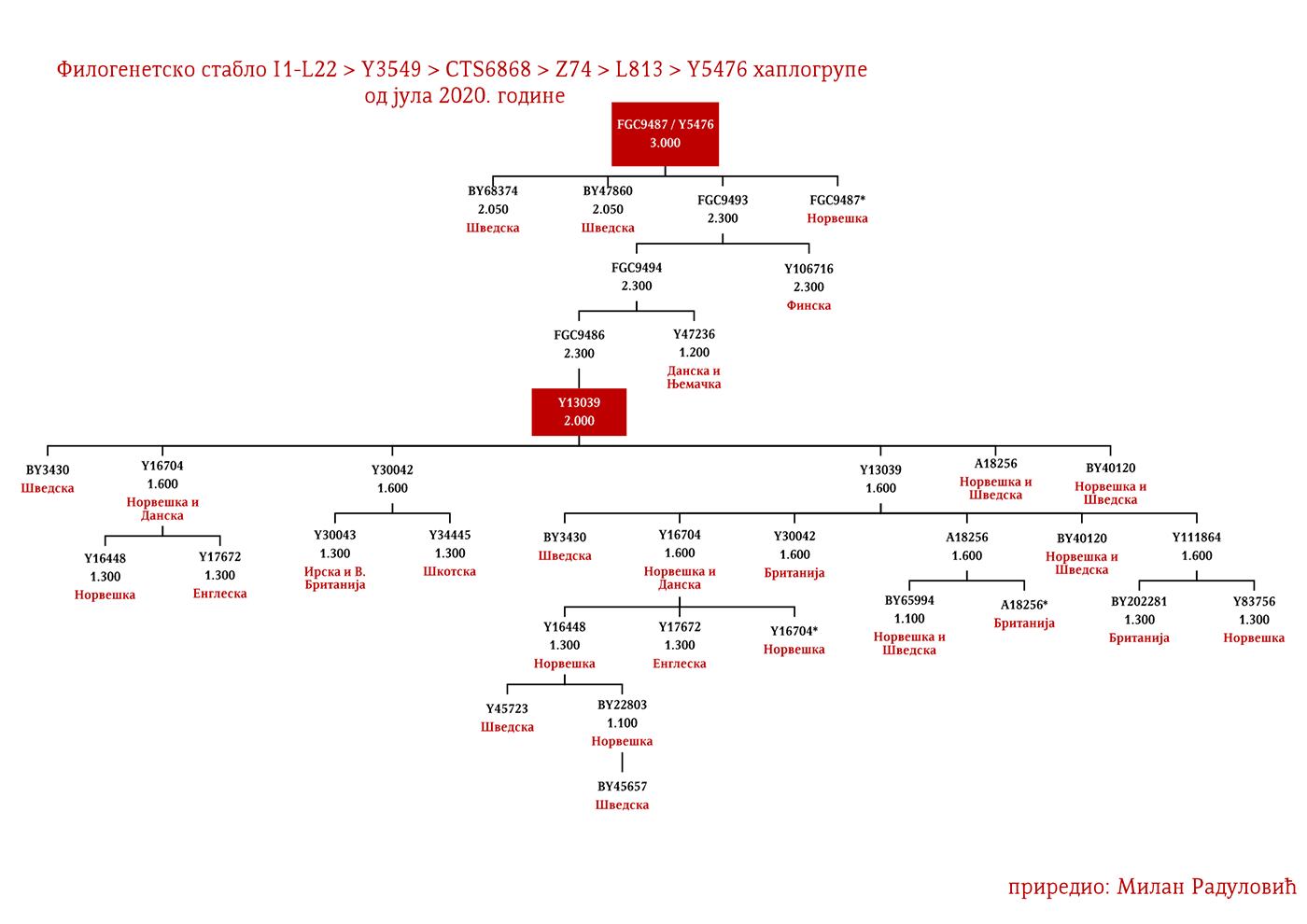
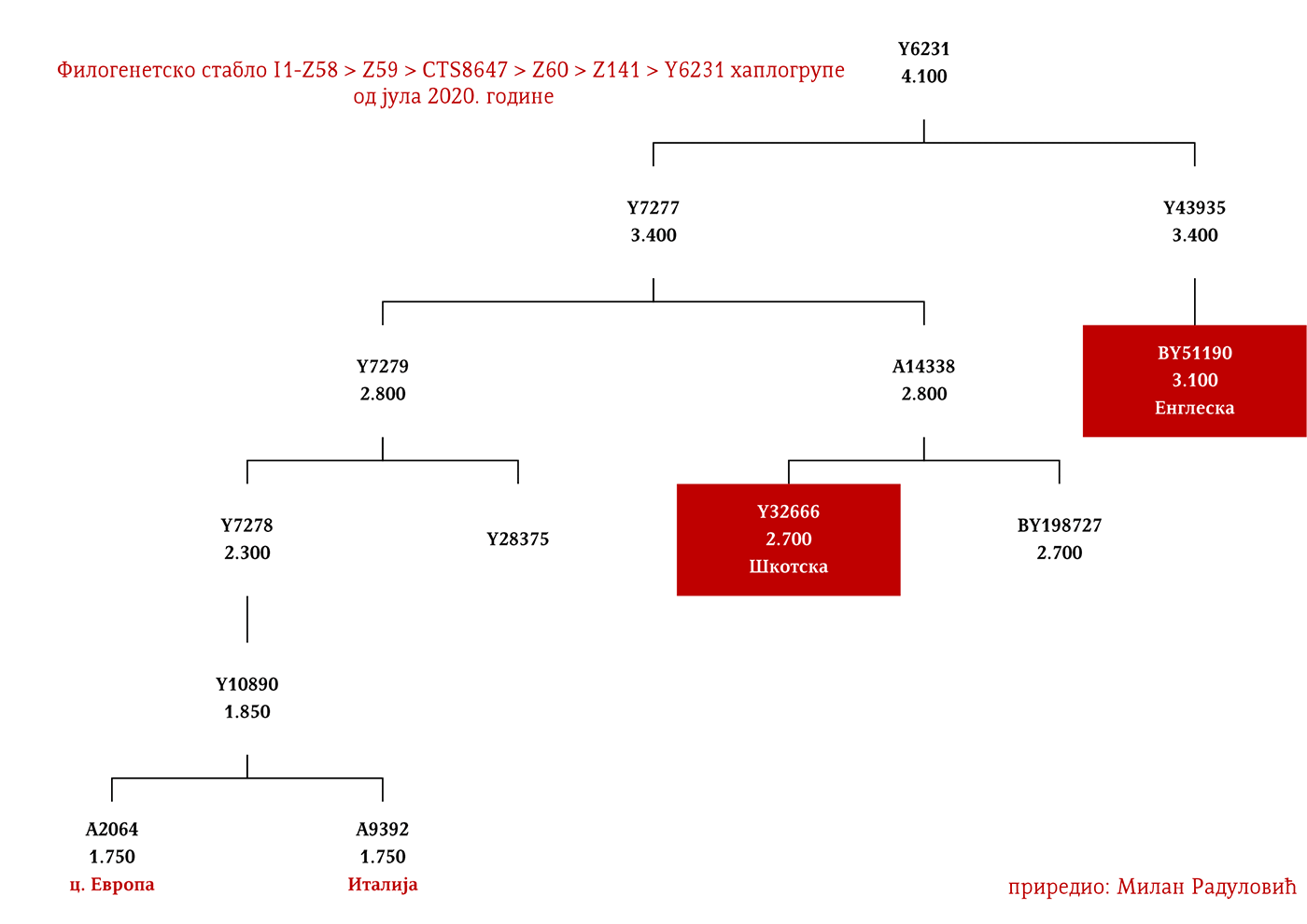
Могуће варјашке линије на основу распрострањености, археогенетике (у тексту као VK), исходишта и старости грана, биле би следеће: I1-L22>Y21972, заступљена у Русији и Шведској; I1-L22>Y31081, заступљена у Украјини, Русији и Шведској; I1-L22>CTS2208*, Русија, VK220; I1-L22>L813>Y79653, Русија, Шведска и Норвешка; I1-L22>CTS2208>FGC12562, Турска; I1-L22>Z26344>FT100268, Русија и Финска; I1-L22>L813>FGC17580, нордијске земље и Русија; I1-L22>P109>FGC21612, нордијске земље, Русија и Енглеска; I1-L22>CTS5350>BY65672, Y22486, Скандинавија и Русија; I1-L22>Z2338, I1-L22>Y21959, I1-L22>CTS2208, I1-L22>L813>Y19207, I1-L22>Y5473, I1-L22>Y22024, I1-Z58>BY61100 – редом VK223, VK20, VK220, VK14, VK221, VK219, VK539.
ГОТИ
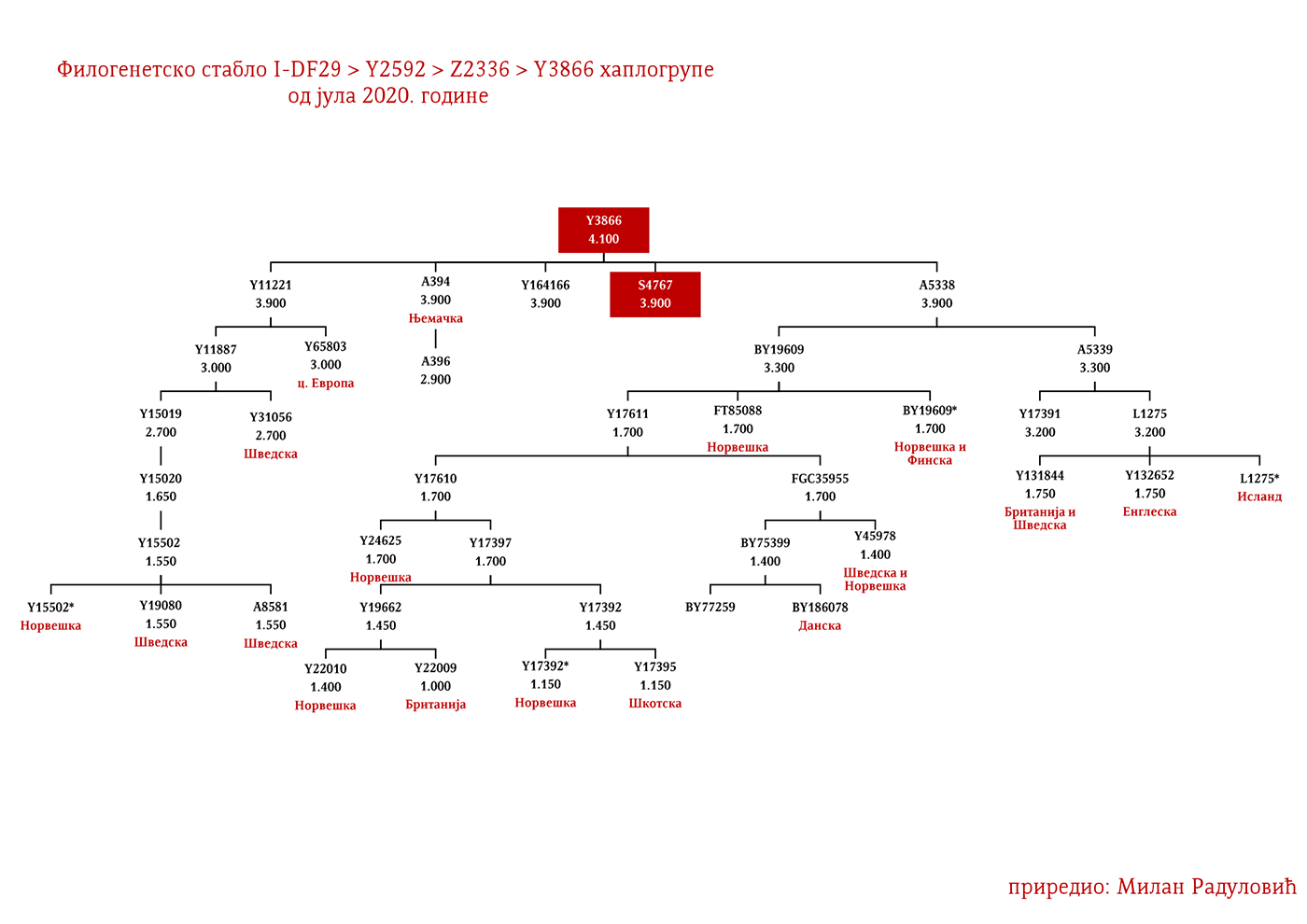
На основу истих горе поменутих параметара Готи би били I1-M227 и следеће I1 линије: I1-Z58>S6402; I1-Z58>Y21391 дио; I1-Z58>A6867; I1-Z58>Z60>Z140>S4700; I1-Z63>S2078>L1242 дио; I1-Z63>S2078>Y16435; I1-Y2592>Y65803; I1-Z63>S2078>Y78357. Наравно ово није коначна оцјена, било је и других грана но за ове са већом сигурношћу претпостављамо.
ВИЗИГОТИ
Визиготске линије би могле бити закључујући према горе поменутом методу: I1-Z63>S2078; I1-Z63>S2078>Y7666; I1-Z63>S2078>Y18660; I1-Z63>S2078>Y59728; I1-Y2592>Y164166; I1-Z58>Z60>Z140>Y29667; I1-Z58>Y7043.
ДРУГИ СТАРИ НАРОДИ
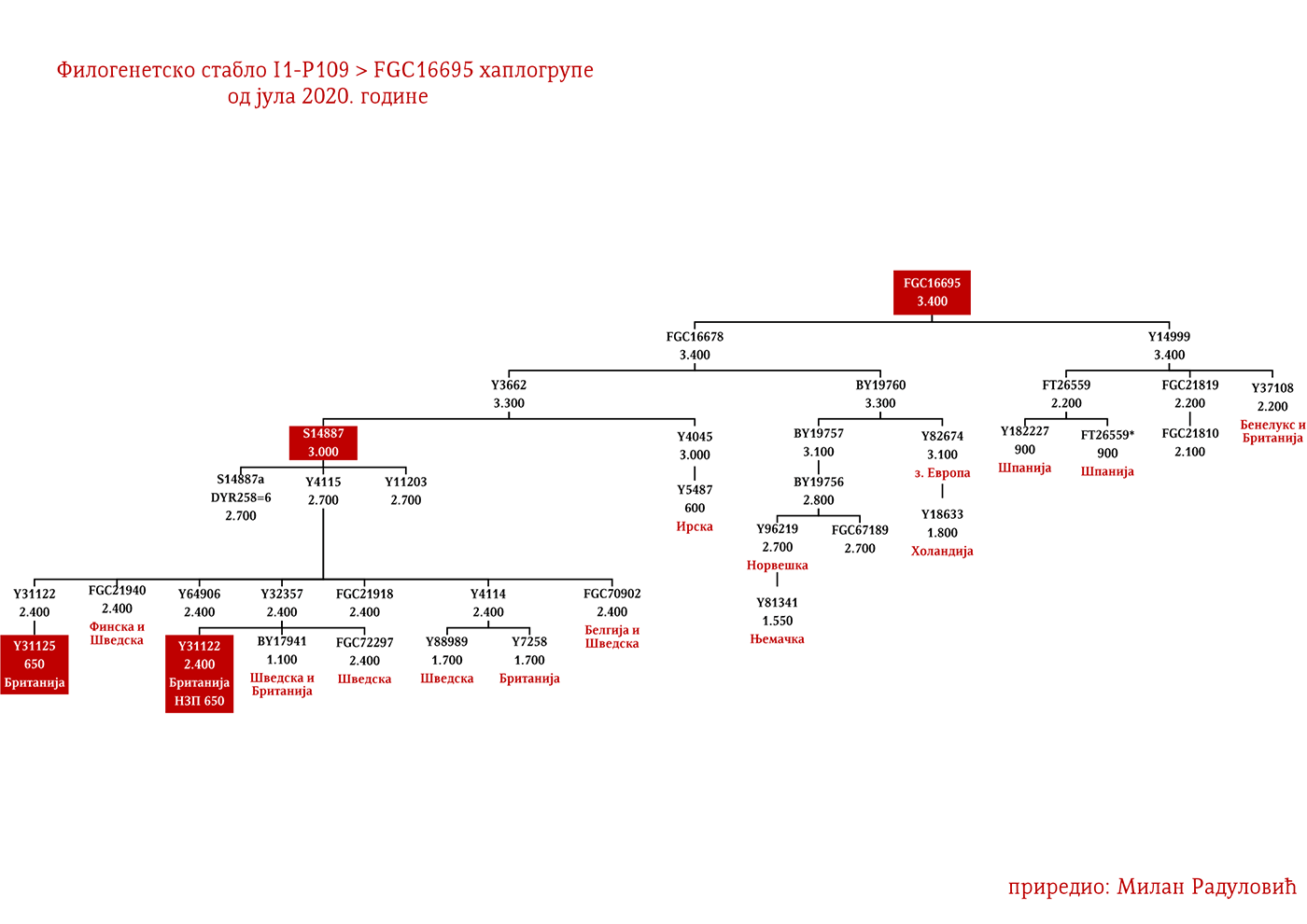
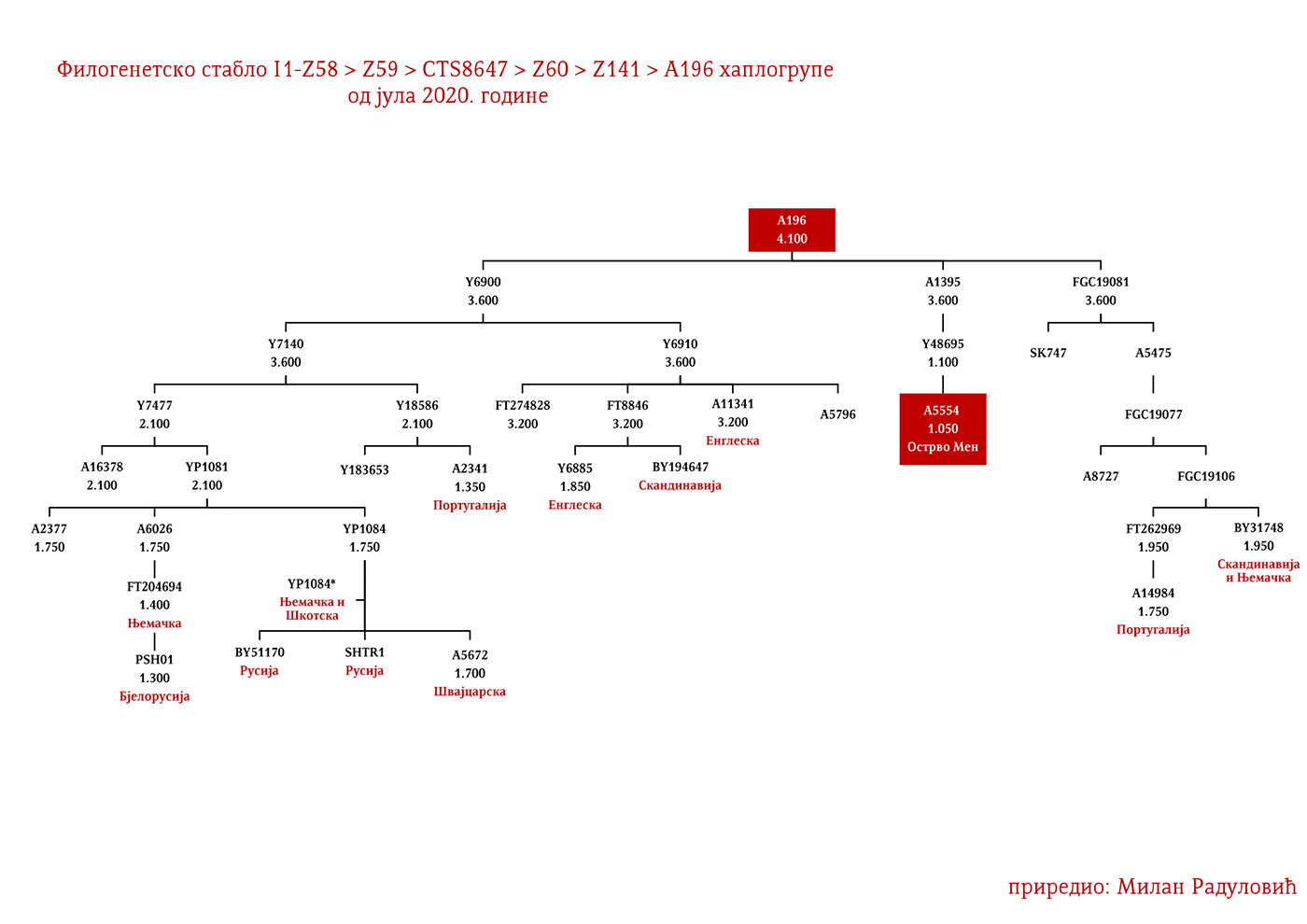
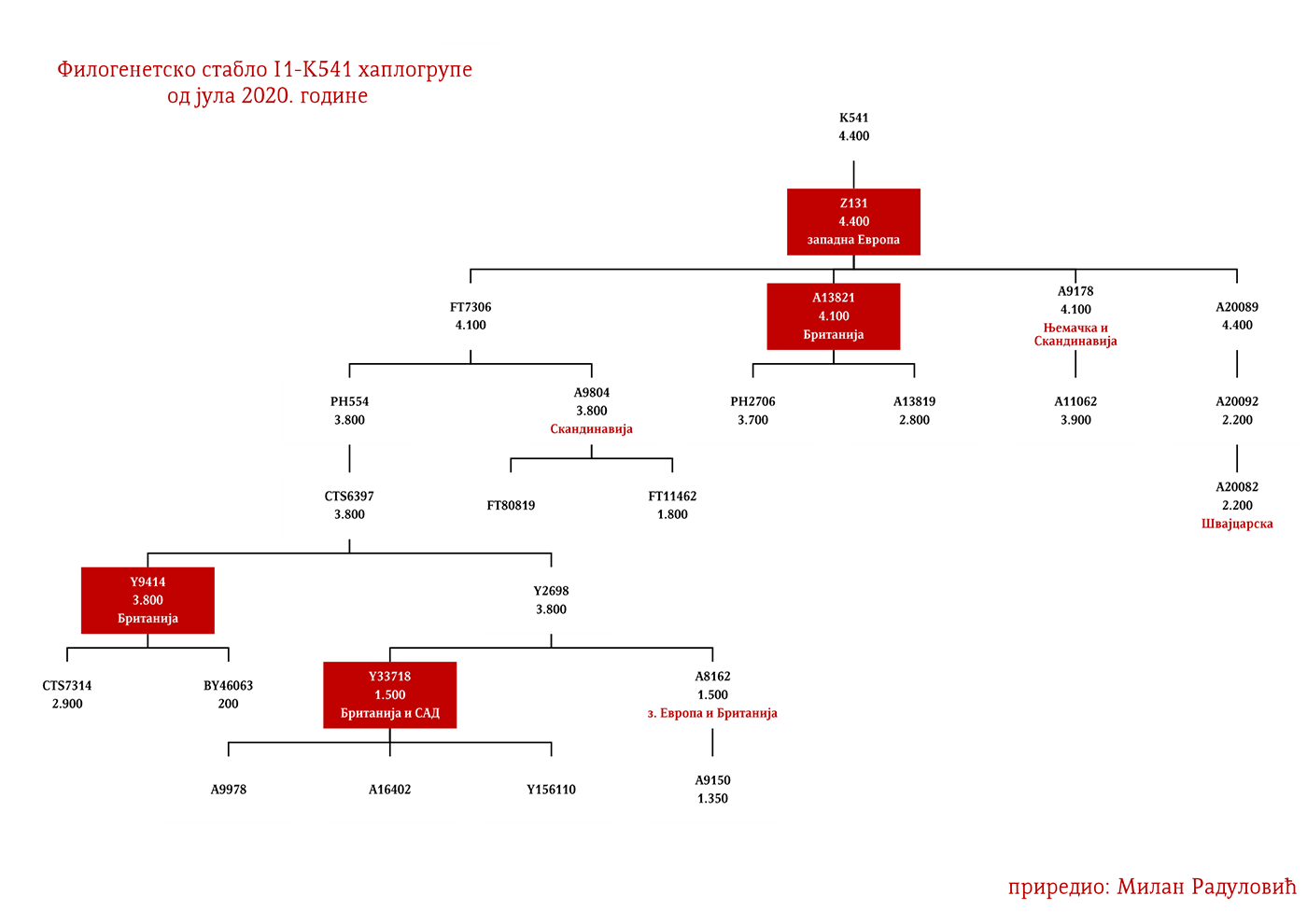
Западни германски народи би махом припадали Z58 и Z63 линијама. Англосаксонске и фризијске гране треба тражити у оним подгранама који се срећу у Енглеској и Бенелуксу, а које имају већу старост од 2000 година. За Лангобарде имамо археогенетску потврду да је једна њихова линија била и I1-P109>FGC21810, узорак SZ45 (Amorim, et al., 2018; Рајевац, 2020).
СРПСКИ НАРОД
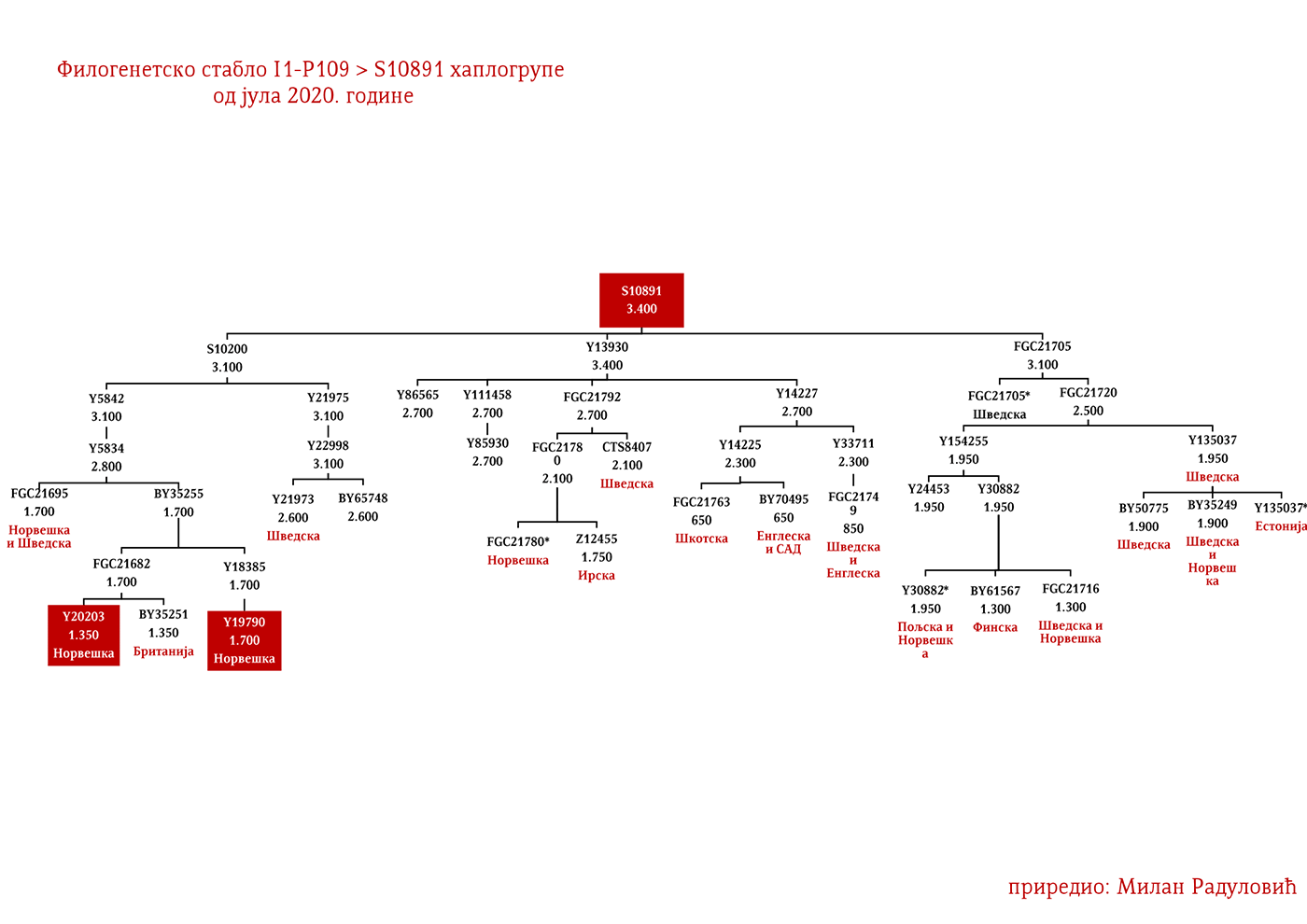
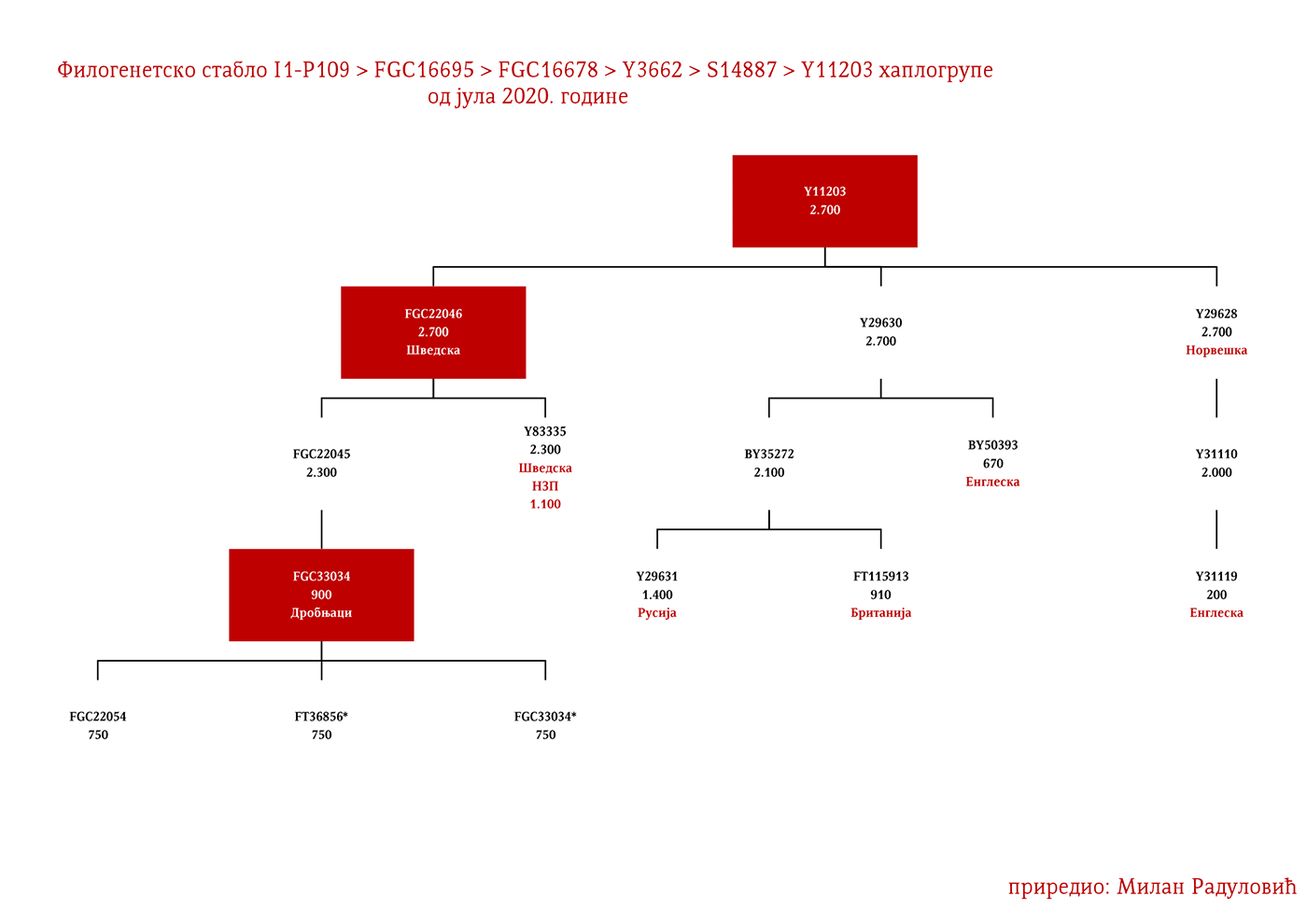
Код Срба најбројни генетички I1 родови су Дробњаци I1-P109>FGC22045 и Мацуре I1-Z63>Y16434, а слиједе их I1-Z63>Y13946>PH220 који су расути на широком подручју иземеђу Крајине на западу и западне Србије на истоку; I1-P109>Y13930 се простиру у динарској зони, а исходе из источне Херцеговине; I1-M227>A11380>BY169301 расути су од Крајине до Шумадије, а доводе се у везу са Готима. Присутне су наравно и друге гране, но ово би биле бројније и генетички одређеније.
ДРОБЊАЦИ I1-P109>FGC16695>FGC16678>Y3662>S14887>Y11203>FGC22046>FGC22045
Први помен Дробњака је из 1354. године када су браћа Дабижив, Богоје и Хран, унуци Дробњакови, подигли цркву у Ваганешу (Јиречек, 1923, стр. 54). О њиховом ранијем поријеклу има више теорија базираних што на предањима што на конструкцијама, стога на овај дио старијег поријекла одговор даје генетичка генеалогија. Према резултатима до којих смо дошли испитивањем Y- ДНК, а који су јавно познати захваљујући Српском ДНК пројекту годинама уназад, Дробњаци припадају I1-P109>FGC22045 хаплогрупи. Од најближих рођака изван Балканског полуострва имају Швеђане и друге нордијске народе. По методологији старијег груписања истраживача, њихов хаплотип сврстава се у кластер познат под називом нордијски. До времена најближег заједничког претка свих тестираних I1-FGC22045 мушкараца свега је 700-800 година (YFull, 2020, pp. 8.06.01-8.09.00 ). Дакле, заједнички предак живио је негдје у 13. вијеку. Изван простора матичне Старе Херцеговине и подручја земаља у којима Срби живе, њихове блиске сроднике срећемо у С. Македонији и на југу Албаније те код тестираног Румуна (вјероватно скорије асимилована линија).
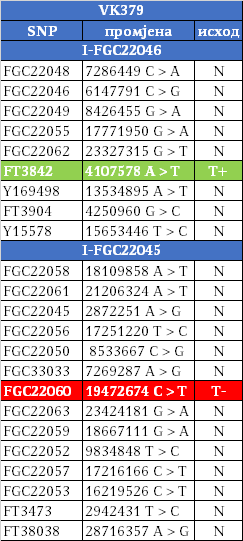
Како су им најближи Швеђани и други Нордијци, те како изостају у резултатима других европских популација и како дјеле заједничког претка у оквиру 2200 година са недавно откривеним Викингом из великог археогенетичког истраживања, VK379 из Шведске, овим потврђујемо писање ранијих српских истраживача Синише Јерковића (Јерковић, 2017), Јовице Кртинића (Кртинић, 2020) и других који су писали о викиншком/норманском поријеклу наших Дробњака. Дакле, свако повезивање FGC22045 са источногерманским народима за сада нема упоришта у резултатима Y-ДНК.
Контакти Нормана и Срба нису историјска непознаница. Скиличин Настављач почетком XII века бележи податак да је Бодинова војска у Бици код Приштине 1072. године заробила византијског војсковођу Лонгивардопула (грч. Λογγιβαρδόπουλος). Заробљени војсковођа беше послат кнезу Михаилу у Зету, који се тако омили кнезу, да му не само слободу врати, већ примивши од њега веру, да ће убудуће њему служити, даде му своју кћерку за супругу. Лонгивардопул добави својих земљака у помоћ, те како бележи Скиличин Настављач, беше састављена нова војска од српских и норманских чета, која под вођством Лонгивардопула би упућена у помоћ Бодину у борби против Византинаца (Рачки, 1873).
Занимљиво, краљ Михаило ће 1081. године и свог сина Константина Бодина оженити Норманком Јаквинтом Баријском. Подсећања ради, исте године су се Нормани под вођством Роберта Гвискара с 16000 ратника искрцали у Драч, гдје су се сукобили са Ромејима, а гдје су Срби предвођени Бодином избегли директно учешће у бици гдје их је позвао ромејски цар (Живковић, 2006). Да ли је ова група ратника остала на простору српских земаља није познато, као што није познато ни шта се на крају десило с норманско-српском војском коју је предводио Лонгивардопул, али је евидентно да је један мушкарац имао оснивачки ефекат неких 100 година касније од ових дешавања.
По народном сјећању које су записали бројни истраживачи, Дробњаци су се по доласку на Језера сукобили са старосједилачким племеном Крича. Борбе између њих биле су дуготрајне и исцрпљујуће. Како су Кричи дали жилав отпор, Дробњацима су у помоћ прискочила друга околна племена, те су Криче потисли према планини Кучајевици гдје су изазвали помор међу њима. Неко вријеме након тога Кричи су се искупили у Фочи и преко Пиве од стране Дурмитора ударили на Дробњаке, али су их ови дочекали. Борбе између Дробњака и Крича биле су дуготрајне и исцрпљујуће, а крајњи резултат борби био је пребацивање Крича на десну обалу Таре (Лубурић, 1930). Народно предање приповједа да је на дробњачком земљишту почела борба против Грка, која је кулминирала у доба Стефана Немање.
Током средњег вијека били су у влашком статусу, а према њиховом предању учествовали су у сукобу Николе Алтомановића и кнеза Лазара Хребељановића на кнежевој страни, предводио их је војвода Милеша Мандић након чега их је кнез Лазар даривао повластицама; у Косовском боју предводио их је Ђурјан Косовчић (Лубурић, 1930, стр. 22, 95).
Ископавањем посмртних остатака из гробнице кнеза Николе Рашковића Дробњака, властелина из 15. вијека, у Церници 2018. године и обрадом његовог Y-ДНК профила непобитно смо утврдили чињеницу да се ради о припаднику дробњачког племена. Но, о кнезу Николи биће више речи у књизи „Генетичко порекло Срба Старе Херцеговине”.
Током отоманске окупације, миграциони правци Дробњака ишли су махом у смјеру других динарских крајева, тако их срећемо заједно са Кричима и Пивљанима у Крајини. Снажно жаришно присуство рода у Полимљу не чуди јер је оно наслоњено на матичну регију, док је јако присуство на Романији последица миграција из Дробњака током бурних година 18. и 19. вијека и нешто мање оних са краја 17. вијека.
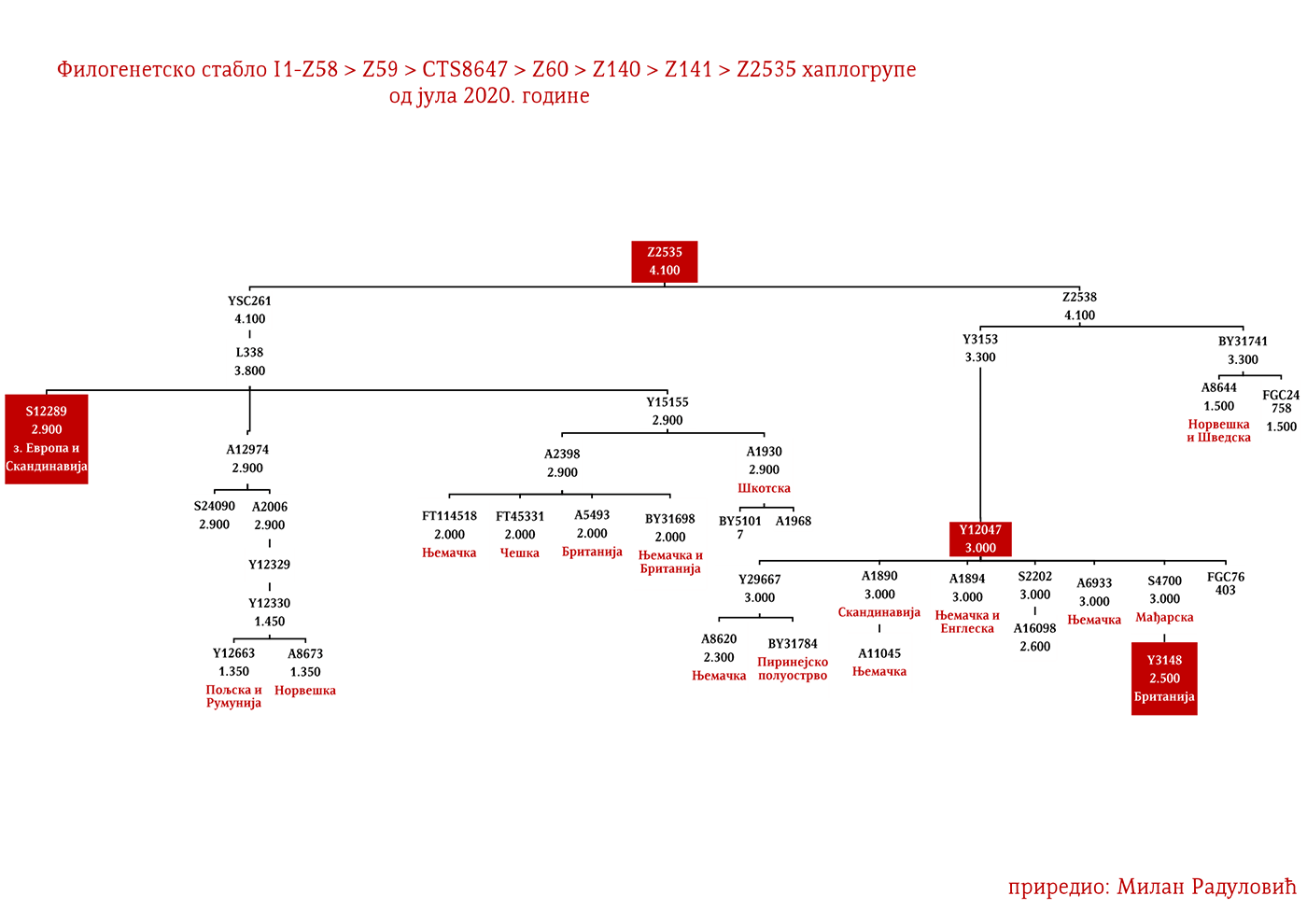
МАЦУРЕ I1-Z63>FGC81364>S2078>S2077>Y16435>Y16437>Y16434
Мацуре су према усменој традицији стари предсловенски народ са простора данашње Црне Горе, из њеног источног дијела, а који је живио на подручју данашњих Пипера, Братоножића, Морачана, Ровчана и Васојевића. Остали су топоними као свједочанство о овом племену, и то су између осталог, пашњак Мацуровине изнад села Лијешње, дио шуме Кунак у насељу Церовица са називом Мацур бријег; бројна стара гробља у селима Трмању, Лијешњу, Вељем Дубоком, Сретешкој Гори, Међуријечју, Церовици те у Горњим Ровцима, где се два гробља називају мацурским, али и у бројним другим местима (Вујанић, Јерковић, Кртинић, & Маринковић, 2014). Како су сви топоними Мацура везани за висоравни и планинске предјеле, аналогно закључујемо да се радило о становништву које се претежно бавило сточарством, односно које је касније током средњег вијека долазило из влашког сталежа.
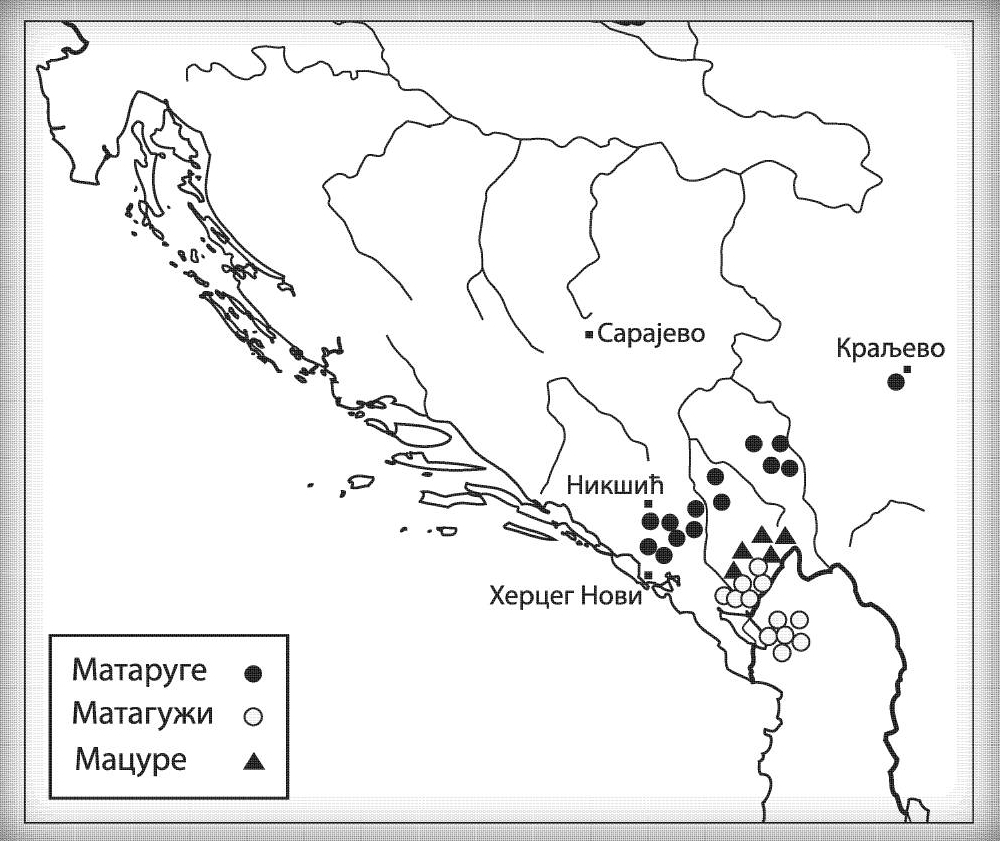
Као разлог наглог нестанка овог племена наводе се оштре зиме, гладне године и њихови међусобни сукоби, те је тако у Црној Гори остала узречица „поклали се к’о Мацуре“ или „побише се к’о Мацуре“ (Вујанић, Јерковић, Кртинић, & Маринковић, 2014, стр. 44). Путем Y-ДНК анализе носилаца презимена Мацура из Крајине и њима сродних братстава из Брда у Црној Гори (в. Табелу Српског ДНК пројекта), утврдили смо прапоријекло овог старог племена, а оно је у најкраћем речено германско и припада хаплогрупи I1- Y16434. Прегледом цјелокупне предачке Y16435 хаплогрупе, највјероватније се ради о линији проистеклој од источногерманских народа.
ДРУГИ СРПСКИ I1 РОДОВИ
Осим дробњачке I1-P109 подгране, на нашим просторима срећемо још најмање двије, а то су Y13930 и Y5621. Хаплогрупу I1-Y13930 срећемо у динарској зони и њено матично подручје је источна Херцеговина; док је Y5621 присутна у околини Врања и вриједи поменути да овој грани припадају два викиншка узорка VK70 и VK281.
Род који за сада није именован а који припада I1-Z63>Y13946>Y51867>PH220 хаплогрупи расут је широм српског етничког простора, од Расине до Босанске Крајине. Даље рођаке ван нашег простора срећемо у Бугарској на удаљености од око 1200 година, а паралелна братска грана Y51867>S22866 присутна је у Швајцарској и Њемачкој, док је заједнички предак Y51867 гране живио прије 2300 година. Равномјернија распрострањеност нам говори у прилог раном учествовању у етногенези Срба, док у комбинацији са претходно наведеним удаљеностима са другим Y51867 мушкарцима излачимо закључак о предсловенском поријеклу рода, односно врло вјероватно се ради о потомцима германских племена која су продрла на Балкан. Наравно могуће су и друге опције досељавања у предсловенском времену, но истакли смо ову вјероватнију.
Још један род који је бројан а није именован, припада подграни I1-M227>A11380>BY169301 и расут је широм српског етничког простора, што нам говори о њиховом раном учествовању у етногенези Срба. Изван Балкана, у оквиру од 2500 година срећемо њихове сроднике у Бјелорусији и Аустрији. Могуће је да су готског поријекла.
Присутне су и друге линије код нас, али како нису нарочито заступљене и како нису довољно профилисане о њима не можемо нешто подробније писати и извлачити закључке.
ПОЗНАТЕ ЛИЧНОСТИ

БИРГЕР ЈАРЛ
Био је јарл Шведске (1248-1266) и зет шведског краља Ерика XI. Долазио је из породице Фолкунга и основао је династију Бјалбо. Представљао је једног од највећих шведских државника средњег вијека. Тестирањем његових посмртних остатака и његовог сина установљено је да припадају хаплогрупи I1 (Malmström, et al., Finding the founder of Stockholm, 2012).
НИКОЛА РАШКОВИЋ ДРОБЊАК
Српски је властелин са подручја Гацка који се помиње у дубровачким списима 1453. године. Током ископавања на локалитету под Маковим валом у Церници крај Гацка 2018. године, уз темеље цркве-задужбине кнегиње Јелене Лазаревић налази се гробно мјесто кнеза Николе чији су узорци тестирани у лабораторији Биолошког факултета из Београда. Ископавање је водио Музеј Херцеговине у Требињу, док је Друштво српских родословаца „Порекло“ било иницијатор истраживачког подухвата. Према резултатима које је обрадио тим Српског ДНК пројекта, кнез Никола Рашковић припада хаплогрупи I1-P109>FGC22045 и роду Дробњака.

ДИНАСТИЈА ГРИМАЛДИ
Династија Грималди је италијанског поријекла и владала је у периоду од 1297. године до 1731. године, када је умро посљедњи члан ове породице. Након смрти посљедњег члана династије Грималди на власт долази династија Матињон-Грималди која је владала до 1949. године. Монаком данас влада династија Полињак-Грималди. Припадају хаплогрупи I1-P109 (Grimaldi – Family Tree DNA, 2020).
ПОРОДИЦА ЛИ
Према подацима са пројекта Ли презимењака (Lee Surname DNA Research Project, 2020, p. Subgroup 21: Lees of VA (Richard Lee m. Ann Constable), позната породица Ли из Вирџиније и Мериленда од којих су били Ричард Први Ли, највећи велепосједник у САД-у и чувени јужњачки генерал Роберт Ли припада I1-P109>S10891>Y13930>Y14227>Y14225>BY70495.

АЛЕКСАНДЕР ХАМИЛТОН
Александер Хамилтон (1757-1804) је један од „Очева оснивача САД“. Био је амерички политичар, економиста и први секретар финансија САД. Захваљујући његовој замисли је сазвана Филаделфијска уставотворна скупштина, а био је и коаутор Федералистичких списа, дјела које се сматра главним извором за тумачење Устава САД. Према резултатима са Хамилтон ДНК пројекта (HAMILTON SURNAME DNA RESULTS AND DISCUSSION, 2020, pp. Group I1-5), Александар је припадник I-Z59>CTS8647>Z60>Z140>Z141>Y15150>Y48678 хаплогрупе.
ЕНДРУ ЏЕКСОН
Џексон Ендру Џексон (1767-1845) био је амерички предсједник и оснивач данашње Демократске странке, такође је био и војсковођа. Познат је по томе што је био 7. предсједник САД. Према анализи генеалогије Хеја (Hay, Thread: President Andrew Jackson belonged to haplogroup I1, 2014) и резултатима са пројекта Џексона (The Jackson Project, 2014), он је припадао I1 хаплогрупи.

ЛАВ НИКОЛАЈЕВИЧ ТОЛСТОЈ
Лав Толстој (1828-1910) био је руски гроф и писац, један од великана свјетске књижевности и један од највећих реалиста тога доба. Познат је по своја два највећа дјела, „Ана Карењина” и „Рат и мир”. Тестирањем његовог потомка Петра Толстоја утврђено је да припада хаплогрупи I1 (Hay, Origins and history of Haplogroup I1 (Y-DNA), 2016).
ДРАГОМИР БОЈАНИЋ ГИДРА
Драгомир Бојанић-Гидра (1933-1993) био је српски позоришни, телевизијски и филмски глумац. Глумио је у више од сто улога у филмовима и ТВ серијама, а улога Жике Павловића у филмском серијалу „Луде године” остала је као култна. Шездесетих година прошлог вијека у Италији снимио је десетак вестерн и акционих филмова. За улогу у филму Балада о револверашу (1967) режисера А. Калтабијана добио је и награду. Припада хаплогрупи I1-P109>FGC22045 и роду Дробњака.








-270x270.jpg)



















3. децембар 2020. у 12:44
Неко,тамо далеко...
Нисам генетичар али једна ствар ми није јасна.По ком критеријуму је И1 старија од И2 хаплогрупе? Тј. зашто је западна грана редни број 1 а источна редни број 2? Процјена настанка И1 је прије 27 500 година.Информација из вашег текста.Оквирно 25 500 прије Христа.На званичној страници Википедије процијењена старост,тј настанак И2 је између 26 000 и 31 000 старе ере.Математика нам говори да је И2 старији минимум 500 година.Као што сам написао,нисам генетичар али на прву лопту нешто не штима.Или сте ви направили грешку па сте умјесто 27 500 прије Христа написали само 27 500 предвидивши и 2020 нове ере или овдје постоји класична дискриминација Истока Европе од стране научних кругова на Западу,тј. истицање своје гране као примарне а мрски источњаци по старом неписаном правилу секундарна категорија? Опет све и да је И1 настала око 27500 године старе ере има ли доказа да је старија од И2 чија могућа старост иде и до 31 000 прије Христа.Да се не своди на оно : њихови институти,њихова наука,њихова правила,њихова категоризација?
Гледајући текст видим и неке занимљивости.
Екстремно мали број тестираних у Норвешкој,Данској и Холандији.На први поглед ми делује да што су људи богатији то их мање занимају овакве ствари…
Албанци су скочили са 2.7 % 2010-те на нешто више од 4 % 2019-те (податак са њиховог ДНК пројекта). Да ли је у питању неко штеловање и манипулација? То је значајан раст.
Изненађујуће низак проценат ове хаплогрупе код балтичких народа.Они редовно истичу себе као “синове Прусије”,тј у секундарној варијанти након свог угро-финског поријекла.Тачно да се то пруско односи на друштвени и културни аспект али реално је било заочекивати значајнији проценат И1 на том простору.Врло су близу германској колевци.југоизападно Њемачка,северозападно Скандинавија .Осим северног краја Естоније који је у близини Финске проценат ове хаплогрупе занемарљив.
Рекао бих да је највеће изненађење управо Србија.Проценат од 8 % на толикој удаљености од матице је И1 заиста фрапантан податак.Ако може неко да ми одговори из администрације сајта или неко ко боље познаје материју каква су даља очекивања тј . неке будуће пројекције заступљености ове хаплогрупе у Србији? .Смањење,повећање или останак на овом проценту? Делује ми на прву лопту као да су Дробњаци и Херцеговци били најзаинтересованији за српски ДНК пројекат…
3. децембар 2020. у 19:13
Бранко Тодоровић
Ове бројке се односе на поједине радове које је аутор користио. То свакако није укупан број тестираних појединаца из неке земље. Тако се могло узети и неко истраживање за Србију, БиХ или Црну Гору, и написати да је проценат узет на основу узорка од 180 људи. Такође, када је реч о уделу хаплогрупе код разних народа, треба имати на уму да нису сви једнако бројни, као и да разновсрност подграна није свуда једнака. Код нас је висок проценат, али је мали број припадника ове хаплогрупе, као што је мали и број њених подграна, за разлику од скандинавских земаља или Немачке, где је њихов број вишеструко већи, по оба параметра.
5. децембар 2020. у 00:26
Dubrovski
Uz svo duzno poštovanje za uloženo u ovu analizu, smatram da je prepostavka da su Drobnjaci Normani zaista smesna. Sami navodite kako je verovatno jedan od tih sto su dosli da pomognu Bodinu posle ostavio svoje tragove. I eto tako vec njegov unuk napravi brojno i mocno srpsko brdjansko pleme. Koi je će to Švedjo ??najamnik da se skrasi u nasim gudurama??? Dajte molim vas.
Normani su uglavnom bili Danci. Veća je verovatnoća da je to gotska grupa, mada i to teško , jer te grupe nema u Španiji , Italiji, Krimu, gde su oni otišli i ostali, sigurno u vecem broju nego na Balkan i sa vise motiva da odu tamo no da ostanu odje.
Baltički narodi su u velikoj vecini R1a, tako i , od Nemaca i Poljaka istrebljeni Prusi. Jako je zanimljivo i za veliku sumnju da su istočnogermanska plemena tako ekspanzivna i vojno napredna naprasno nestala i prepustila celu Istočnu Evropu , pa i pola Njemačke miroljubivim Slovenima iz močvara Pripjata, koji su, gle čuda za sto godina okupirali i do dan danas zadržali svoje ime u čitavoj Istočnoj Evropi.
Srdačan pozdrav
5. децембар 2020. у 00:30
Dubrovski
Koja je haplogrupa Varjaga? R1b ili I1? slabo su nešto ostavili traga ti Švedjani po Ukrajini i Rusiji?
Varjag je slovenski izgovor švedskog narodnog imena SVERIGE
9. децембар 2020. у 12:13
Горан
Народ који је такође дуго времена боравио на Балкану и који веома много утицао на историјска дешавања на југу Европе јесу Готи.Готи су свакако оставили и значајан генетски траг на Балкану собзиром дасу и Визиготи дуго времена боравили у Дакији и Тракији,ишли до Епира а са крајњег југа Визиготи су отишли преко Далмације до Сев.Италије и даље на Запад.Остроготи су боравили у Панонији одакле су отишли у Превалис(Дукља,Зета)одакле одлазе у Италију а затим формирају своју краљевину која обухвата Италију и сву Балканску Далмацију.Њихова краљевина на Балкану је трајала око 50година аи после пораза Острогота од Византинаца,многи Остроготи на Балкану нису тренутно нестали већ су многи остали у брдима Балкана у 5 и 6веку где су дочекали Словене.Неки су били полуроманизовани сточари планинци названи Власи,а неки су у 6,7веку били Словенизовани.У генетском смислу су за Србе и друге Балканске народе веома битни јер су дуго времена били присутни на Балкану скоро три века,били су пореклом из Скандинавије из Готланда и отишли су далеко на исток према Црном Мору типични I1Z63.Још док су били у заједници као Готи пре поделе на Остроготе и Визиготе док су живели на ушћу Висле ту су боравили заједно са Бургундима који су такође пореклом Скандинавци Англи са острва Борхолм.Готи су повели један број Бургунда са собом према реци Одри где сусрећу Вандале.Са Готима иду и Гепиди и у близини Црног мора сусрећу Германске Бастарне и Скире.Такође на реци Одри могли су им се придружити и бројни Ругијци.Тако су Готи као народ дошли на Балкан са већим бројем Германских хаплогрупа подграна водећи са собом припаднике већег броја Источних Германских племена која свако има своју генетску подграну.Тако напр.Готи су асимилирали један број Бургунда још на ушћу Висле,који су већ у 4веку били потпуно интегрисани у Готе и Источне и Западне.Бургунди би могли да буду носиоци I1Z63-51867 и ови Готи-Бургунди су могли ту подграну да донесу иу Бугарску и Македонију,Србију,Херцеговину,Далмацију,тако иу Италију,Шпанију,где су све ишли Готи.Такође Готи су собом водили и групе Вандала,Гепида Ругијаца који су Готима донели I1Z63-16435 које су са Готима донели на Балкан.Готи су собом водили и групе Бастарна и Скире који су Готима пренели I2M227 коју су као Готи донели у Далмацију.Сами Готи као Скандинавци су могли да буду носиоци I1Z63-L22 која је типична за Скандинавце а која је доста бројна у Старој Херцеговини поред других Готских подграна (I1Z63-51867-Ph220),I2M227, I1Z63-16435,што су све имала Германска племена која су асимилирали Готи на свом путу од Скандинавије ка јужној Европи.Ове Готске,,Германске,,хаплогрупу-подграну су доказ дасу многобројни Готи остали и били Словенизовани на Балкану.
7. март 2021. у 17:58
Горан
Многи родови који имају аплогрупа I1Z63 своје порекло везују за Стару Херцеговину,Никшић,Стару Жупу Оногошт и Грахово.Многи родови који су I1Z63 су из Оногошта у средњем веку отишли према Дубровнику,Требињу,Котору,Поповом пољу,Гацку.Скоро сви I1Z63 у Старој Херцеговини могу се повезати са Старим Оногоштом,Никшићком пољем и тврђавом Анагастум.Ову тврђаву у 5.веку су подигли и у њој иу пољу око ње,живели Остроготи.У време досељавања Словена у ширу околину тврђаве Анагастум у 7веку,потомци Острогота из Анагастума остали су у пољима и жупи поред тврђаве,где су се сјединили са досељеним Словенима R1a.Собзиром на висок проценат I1Z63 хаплогупе код родова који су пореклом од Оногошта-Никшића-Риђана,може се закључити даје већи рој Острогота остао у брдима и пољима око Анагастума где су дочекали Словене који су тврђаву назвали Оногошт,и ту формирали своју Жупу.Вероватно да између Острогота и Словена у Оногошту није било никаквих сукоба,већ су Остроготи били добровољно врло брзо Словенизовани и остали да као Словени живе у Жупи.Можемо слободно рећи да су Срби са хаплогупом I1Z63, пореклом од Никшића,потичу од Словенизованих Острогота из Анагастума,тврђаве са границе Превалиса.