



PIŠE: Milan Radulović ([email protected])
Objavljeno: 30.11.2020. | Dopunjeno: 30.11.2020.
Preuzmite besplatno PDF verziju članka.
OPŠTE NAPOMENE
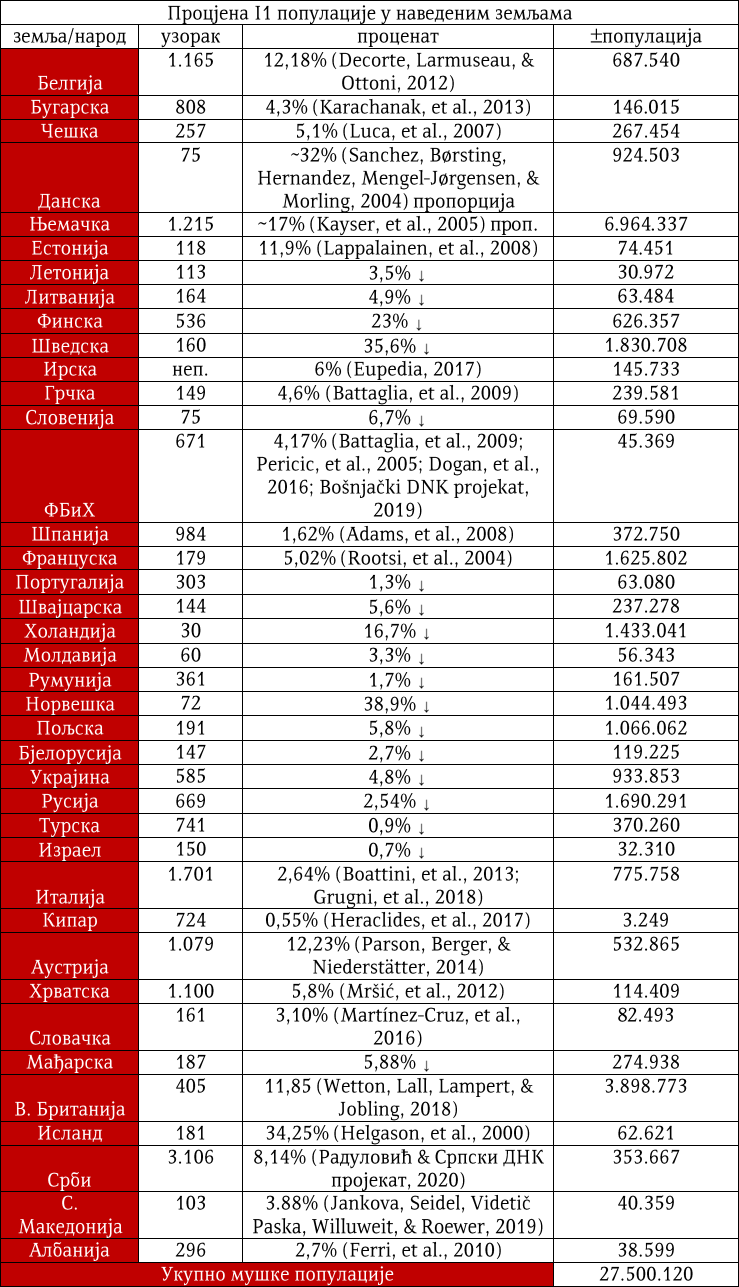
Haplogrupa I1 proistekla je iz predačke haplogrupe I. Ona se od nje odvojila prije 27 500 godina. Zajednički predak svih današnjih pripadnika haplogrupe I1 živio je prije 4 600 godina i to negdje na prostoru centralne Evrope. Haplogrupa I1 može se povezati sa ekspanzijom germanskih naroda. Prema podacima Srpskog DNK projekta, ona je kod Srba zastupljena u procentu od 8,32%.
PORIJEKLO I MIGRACIJE
Haplogrupa I1 nastala je prije 27 500 godina i bila je na ivici opstanka do prije 4 600 godina kada je živio kako je prethodno pomenuto zajednički predak svih današnjih I1 muškaraca (YFull, 2020), moguće na prostoru Danske (Nordtvedt, 2008). Za mjesto nastanka haplogrupe smatra se širi prostor Evrope. Predstavljali su prve lovce sakupljače Evrope zajedno sa pripadnicima I2 haplogrupe, a u neolit su ušli sa Kulturom linearne keramike u kojoj su bili aktivni sudionici. Najstariji uzorci su iz Španije i Švedske. U Španiji su pronađeni uzorci koji su predački u odnosu na I1-M253 mutaciju, BAL001/005 iz gornjeg paolelita datiran u 11 384-10 733 p.n.e. sa lokaliteta Balma Gilanja u Kataloniji (Villalba-Mouco, et al., 2019) i I10899/Car1 datiran u 9700–5500. god. p.n.e. (Olalde, et al., 2019). Širenje po Evropi započinju tokom gvozdenog doba i intenzivno tokom antike i ranog srednjeg vijeka.
MEZOLIT
Tokom mezolita, lovačko-sakupljačkog perioda, I1 muškarci bili su rasprostranjeni od Pirinejskog poluostrva do centralne Evrope, na šta nam ukazuju nalazi drevne DNK. Kada su prvi ljudi kročili na tlo Skandinavije posle povlačenja leda prije nekih 11 700 godina, formirali su i prve kulture. Na malenom ostrvu Karsle, blizu Gotlanda, na lokalitetu Stura Karlse, među lovačko-sakupljačkom zajednicom pronađen je uzorak koji prethodi I1 haplogrupi kod analiziranog muškog skeleta SF11 datiranog u period između 7050-6850. god. p.n.e. (Günther, et al., 2018); dok ostala tri skeleta iz istog rada pripadaju I2 haplogrupi, odnosno uzorak SBj.SG iz Gotlanda datiran između 7050-6650. god. p.n.e. haplogrupe I2a > Y4213; drugi skelet pod šifrom Steigen.SG iz Moleja, Stejgena, Norveška, datiran je između 4050-3850. god. p.n.e., I2a > M423; i treći I2c > S6635, Hum2.SG, 7550-7350. god. p.n.e., takođe iz Norveške, sa lokaliteta Humervikolmen. Ono što je uočljivo iz rada jeste da testirani uzorci sa istoka Skandinavije imaju pretežno zapadne genetičke primjese lovačko-sakupljačkih zajednica, dok uticaj istočnoevropskih lovačko-sakupljačkih zajednica uočavamo u zapadnim djelovima Skandinavije. Pravci migracija istočnih i zapadnih lovačko-sakupljačkih zajednica kao i njihovi udjeli u mezolitskom stanovništvu Skandinavije prikazani su u Prilogu 1. Nalaz SF11 u samom radu nije označen kao prethodnik I1 haplogrupe, zbog lošije pokrivenosti Y-DNK, pa je ovom prilikom pristupljeno analizi sirovog genetičkog materijala radi potvrde pisanja Genetikera (Genetiker, 2018) koji je uočio da se radi o muškarcu sa mutacijom koja je predačka u odnosu na I1 haplogrupu. Nakon obrade sirovih podataka, potvrđujemo da se radi o muškarcu sa mutacijama koje su rođačke u odnosu na I1-M253, a to su sledeće: Z2726 G+, YSC0000301/Z2882 C+, CTS6221 T+, CTS1748 G+, Z2731 A+, CTS6629 A+, CTS5887 C+, Y7398 T+, dok snip M253 nije pokriven laboratorijskom analizom.
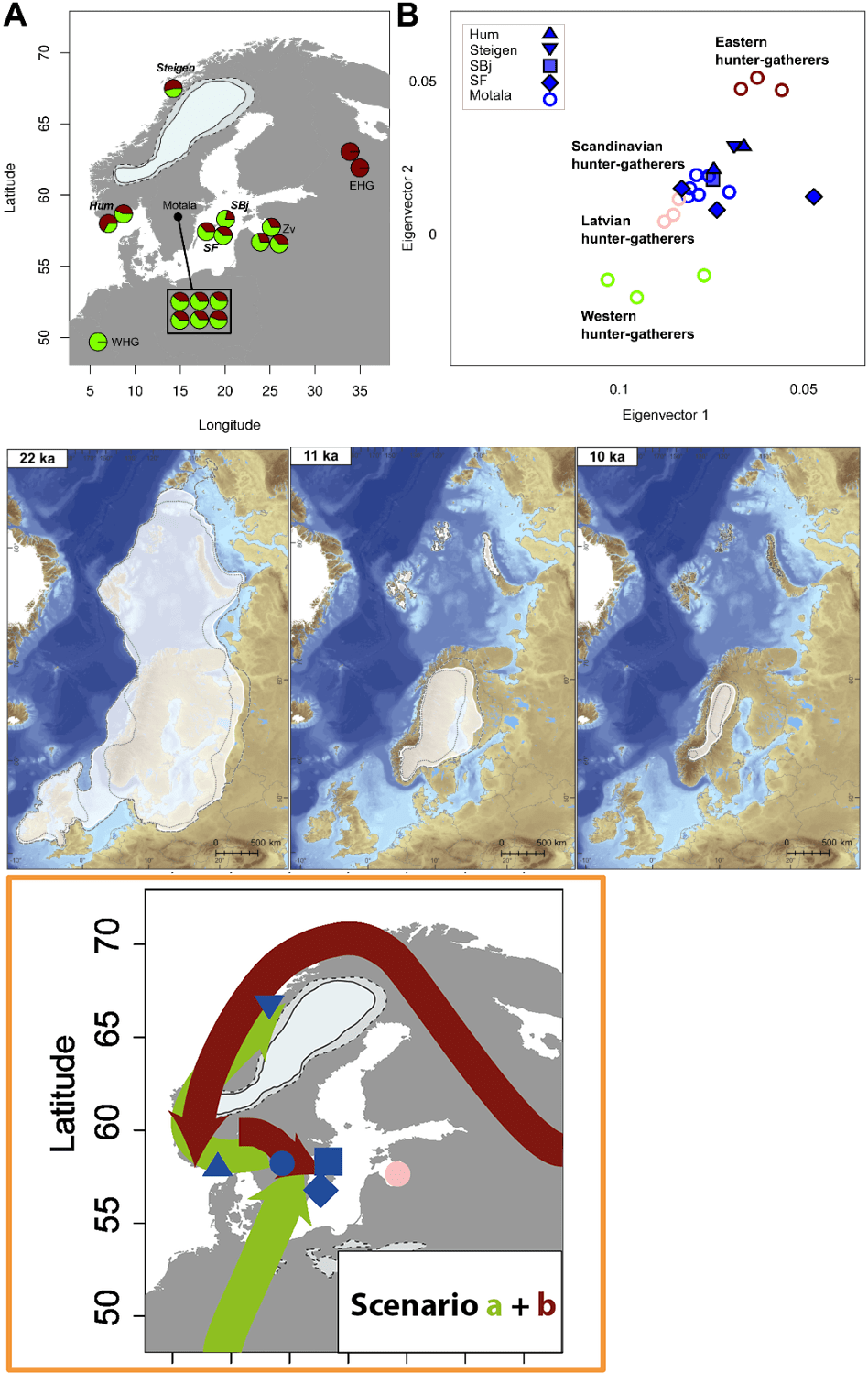
Drugi rad koji se bavio mezolitskom drevnom DNK jeste rad Haka i saradnika iz 2015. godine, koji su analizirali Y-DNK 5 muških skeleta iz Motale u Švedskoj (Haak, et al., 2015). Kod svih je ustanovljena I2 haplogrupa. Uzorak I0012, datiran između 5898-5531 p.n.e, pripada I2c-L596 haplogrupi; I0013, datiran između 5898-5531 p.n.e, pripada haplogrupi I2a>M423; I0015, 5898-5531 p.n.e. je I2a>P37; a I0017 pripada I2a>L1498.</>
Iz priloženih radova zaključujemo da su muškarci sa mutacijama koje su rođačke u odnosu na I1-M253 bili prisutni među prvim stanovnicima Skandinavije s tim da nisu bili reprezentativnije zastupljeni u udjelu mezolitske populacije sjevernih djelova Evrope, koji su dominantno bili I2 haplogrupe. Kako pouzdano znamo da je predak današnje I1 populacije živio prije 4600 godina na području centralne Evrope, o čemu će biti riječi u nastavku rada, zaključujemo da pomenuti mezolitski muškarci sa ranim I1 mutacijama nisu ostavili potomstva među današnjom populacijom, kao ni ranije pomenuti sa područja Pirinejskog poluostrva, dakle radi se o linijama koje su izumrle.
NEOLIT
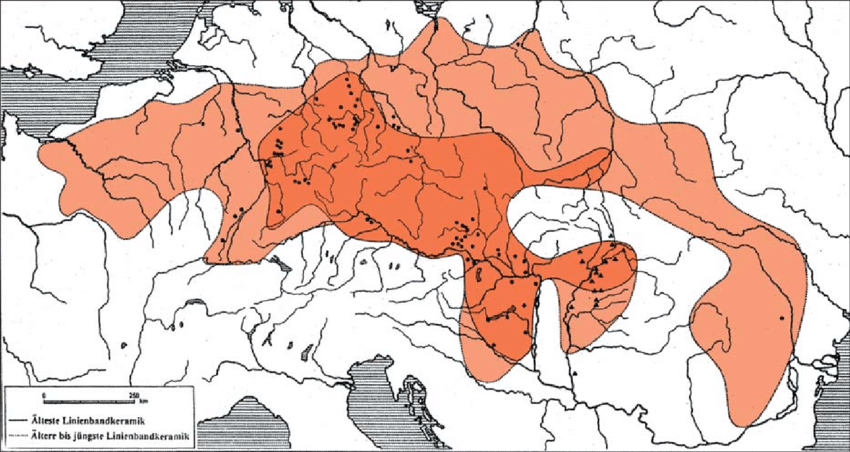
Za neolitsku revoluciju u centralnoj Evropi zaslužna je Kultura linearne keramike (5500-4500. god. p.n.e.) koja je ujedno važna i za današnju I1 populaciju. Razvila se na području Prekodunavske Mađarske gdje je koegzistirala sa južnoevropskom Starčevo-Kereš-Kriš kulturom koja je imala razvojnog uticaja na nju (Banffy, 2000). Zahvatala je Mađarsku, Slovačku, Češku, Poljsku, sjeverni Banat, jugozapadnu Ukrajinu, Njemačku, Holandiju, Belgiju, Francusku, Švajcarsku i Austriju. U Prilogu 2 nalazi se karta na kojoj je prikazana rasprostranjenost kulture. Područje Kulture linearne keramike genetički su pokrile studije Lipsona i saradnika kao i Sidžinji-Naj i saradnika, Nikitina, Matinsona, Gambe i Haka. Kako je u ovoj kulturi pronađen najstariji potvrđeni I1-M253 uzorak, za potrebe rada predočićemo sve analizirane uzorke iz pomenute neolitske kulture radi sticanja sveobuhvatne slike i konteksta u kome su živjeli I1 muškarci. Od 38 analiziranih muških skeleta samo je jedan pripadao I1 haplogrupi dok njih 18 pripada G2 (G2a, 18 i G2, 2) haplogrupi; 6 ih je I2 haplogrupa; 4 su C1a i jedan je CT haplogrupa (bliža grana nije određena); dva uzorka su T1 haplogrupa; haplogrupi F pripadaju 2 uzorka (bliža haplogrupa nije određena), haplogrupi H 2 uzorka, haplogrupi J2a 2 uzorka i jedan uzorak pripada R1b haplogrupi. U Tabeli 1 (v. članak u .pdf verziji) prikazani su svi dosadašnji uzorci Kulture linearne keramike.
Iz priloženog se uočava da su u Kulturi linearne keramike pretežno učestvovale haplogrupe anadolskih farmera G2a, J2a i T1 dok su manje zastupljene I2a, I1 i C1a haplogrupa, svojstvene prvom stanovništvu Evrope, lovačko-sakupljačkim zajednicama koje su se priključile ovoj zajednici zemljoradnika. Dalje, iz ovih rezultata zajedno sa ranije pomenutim mezolitskim uzorcima, kao i iz rezultata drugih studija koje su se ticale evropskih neolitskih kultura, a u kojima nije pronađena I1 haplogrupa, izvlačimo analogni zaključak da su preci današnjih I1 muškaraca bili grupisani na širem području Kulture linearne keramike.
Sledeća neolitska kultura koja bi bila od važnosti jeste Kultura levkastih pehara (4300-2800. god. p.n.e.) koja se prostirala na području današnje Danske, Poljske, Ukrajine, južne Skandinavije i u njenom kasnom periodu do Bjelorusije. Predstavlja prvu zemljoradničku kulturu Skandinavije koja je koegzistirala sa kulturom lovaca sakupljača. Nažalost ona nije bila u širem fokusu istraživača koji su radili analize drevne DNK, te je za sada analizirano svega nekoliko muških skeleta pripadnika KLP-a (Kultura levkastih pehara), njih 6. Jedan pripada IJK haplogrupi (bliža haplogrupa mu nije određena), a evidentiran je pod šifrom I0 802, datiran između 3400-3025. god. p.n.e.; uzorak ros5 je I2-pra-S2742, datiran između 3090-2920. god. p.n.e.; I0 551 pripada G2a-PF3147 haplogrupi, 3400-3025. god. p.n.e.; I6677 je H2 haplogrupa, 3800-3400. god. p.n.e.; I0 807 je I2 haplogrupa, 3970-3710. god. p.n.e.; i I0 559 je R1b-V2219 haplogrupa, datiran između 3654-3527. god. p.n.e. Iako nije pronađena u ovih 6 uzoraka, prisustvo I1 haplogrupe ne može se isključiti iz tri razloga. Prvi je taj što je trenutni uzorak suviše mali, drugo, KLP se naslanja i vremenski i geografski na Kulturu linearne keramike i treći razlog je taj što u genetičkim primjesama testiranih pripadnika Kulture levkastih pehara iz Švedske postoji dominantna, nadpolovična genetička komponenta pripadnika Kulture linearne keramike (pogledati Prilog 3).
Ovde ćemo dodati da je u radu Alentoft i saradnika (Allentoft, et al., 2015), pronađen kasnoneolitski I1 uzorak sa juga Švedske zaveden pod šifrom RISE179, datiran u period 2010-1776. god. p.n.e, što dodatno potvrđuje neolitsko prisustvo I1 muškaraca na jugu Skandinavije. Uzevši priloženo u obzir, sasvim sigurno možemo smatrati da su preci današnjih I1 muškaraca naseljavali pored centralne Evrope i južnu Skandinaviju, već tokom skandinavskog neolita, odnosno od Kulture levkastih pehara. Dakle područje gde su bili grupisani tokom neolita se nalazilo na širem području ove dvije kulture. Kako je zajednički predak današnjih I1 muškaraca živio nekih 2000 godina nakon Kulture linearne keramike, odnosno nekoliko stotina godina posle Kulture levkastih pehara, mi ne možemo sa sigurnošću tvrditi ništa više od gore navedenog. Ostaje nepoznato koji događaji su uticali na strahovito usko grlo ove haplogrupe.
ENEOLIT
Na sjeveru Evrope eneolit počinje nešto posle 2800. godine p.n.e i završava se oko 1700. godine p.n.e, kada nastupa bronzano doba. Kultura vrpčaste keramike (2900-2350. god. p.n.e.) je karakteristična kultura kasnog neolita i eneolita. Glavna je praistorijska indoevropska kultura sjeverne i srednje Evrope, prostirala se od Holandije i Švajcarske na zapadu, pa na istok kroz srednju Evropu do gornjeg toka Volge i srednjeg toka Dnjepra, te na sjever u Skandinaviju.
Pogodna klima, nalik onoj današnjoj sa sjevera Francuske, u Skandinaviji je trajala od 2700-850/650. god. p.n.e, što je doprinjelo naseljavanju pripadnika Kulture vrpčaste keramike (Kane, 2019).
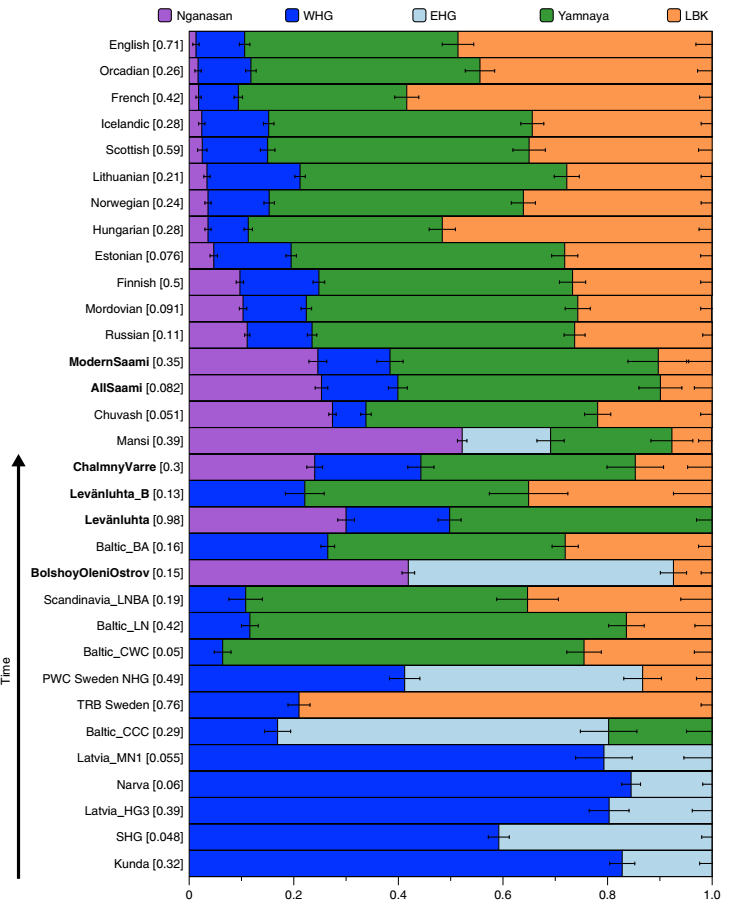
Da bismo stekli cjelovitu sliku prikupili smo sve arheogenetičke radove koji su se bavili ovom kulturom i analizirali Y-DNK njenih pripadnika. U Tabeli 2 (v. članak u .pdf verziji) nalazi se kompletan pregled svih 37 prikupljenih uzoraka. U ovoj kulturi su pretežno bili zastupljeni R1a i I2a muškarci, a manje su zastupljeni pripadnici haplogrupa R1b i G. Muški I1 skelet sa lokalietea Else (Ölljsö) nalazi se pod šifrom oll009 datiran između 1930-1750. god. p.n.e, a prema analizi našeg stručnjaka za drevnu DNK Milana Rajevca, koja je u formi detaljne analize urađena namenski za potrebe ovog članka, oll009 pripada I1a-DF29 haplogrupi (Rajevac, 2020). Drugi interesantan uzorak je već pomenuti RISE179 sa lokaliteta Abekos (Abekås I), koji je takođe I1, sa područja Kulture vrpčaste keramike.
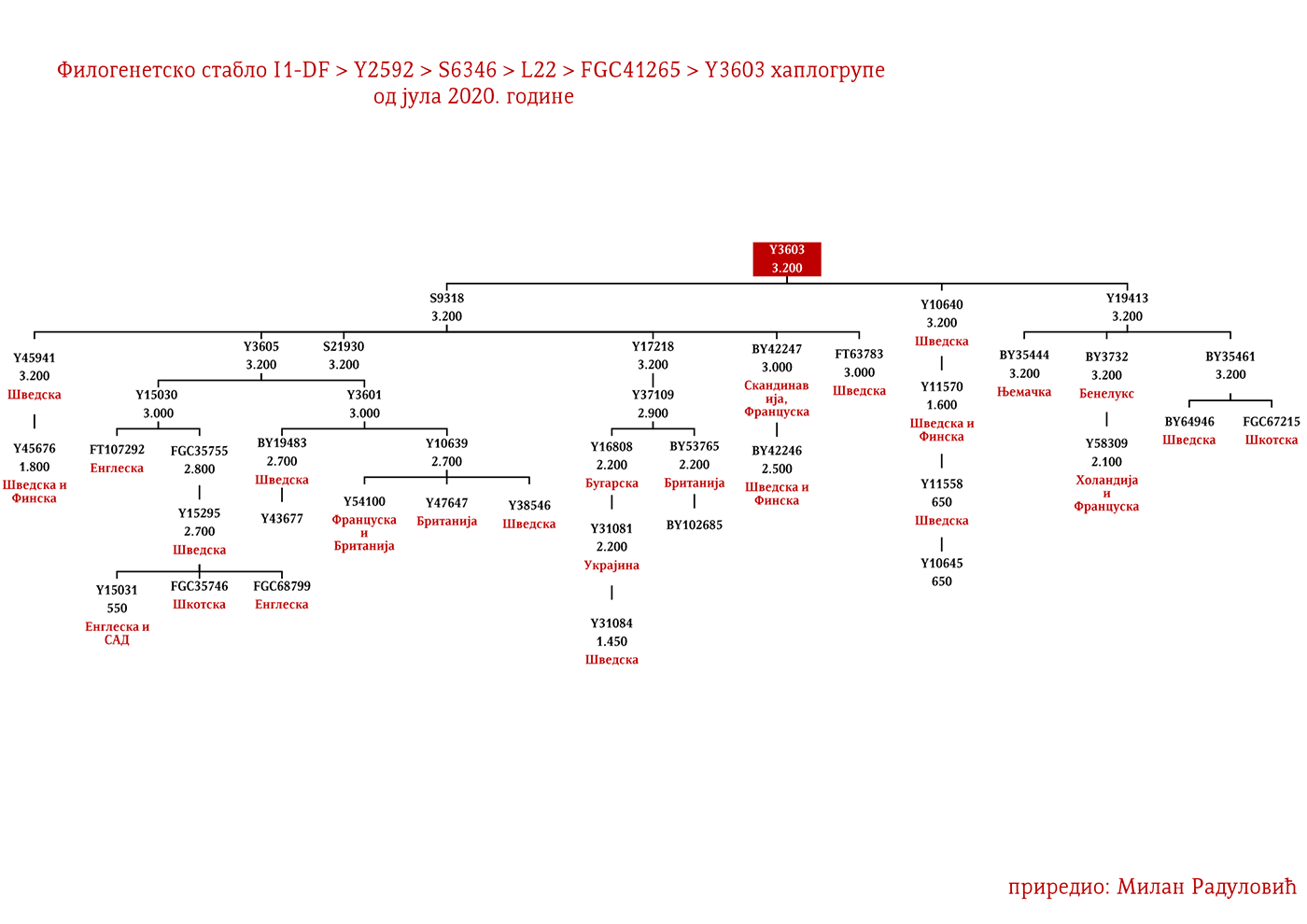
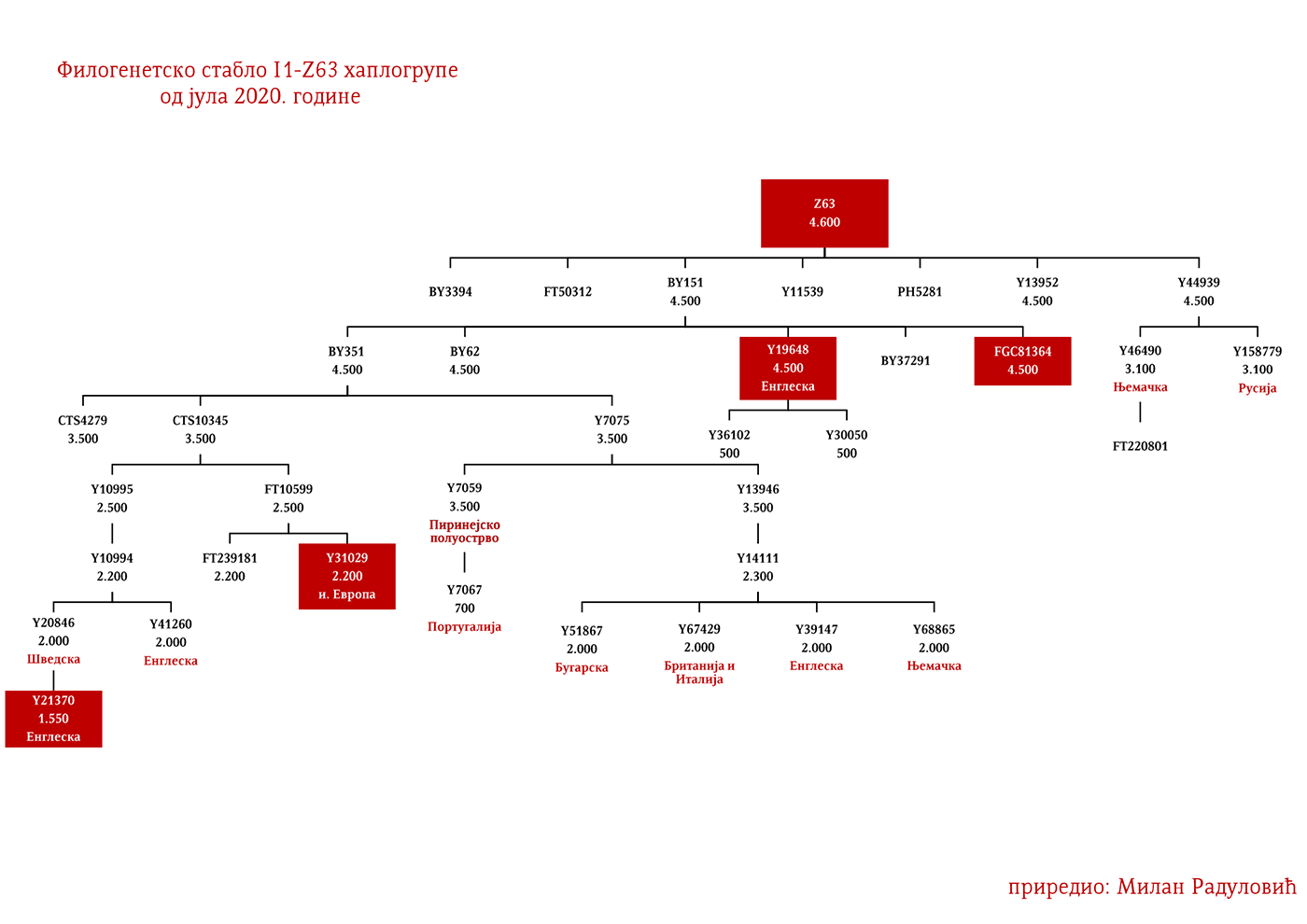
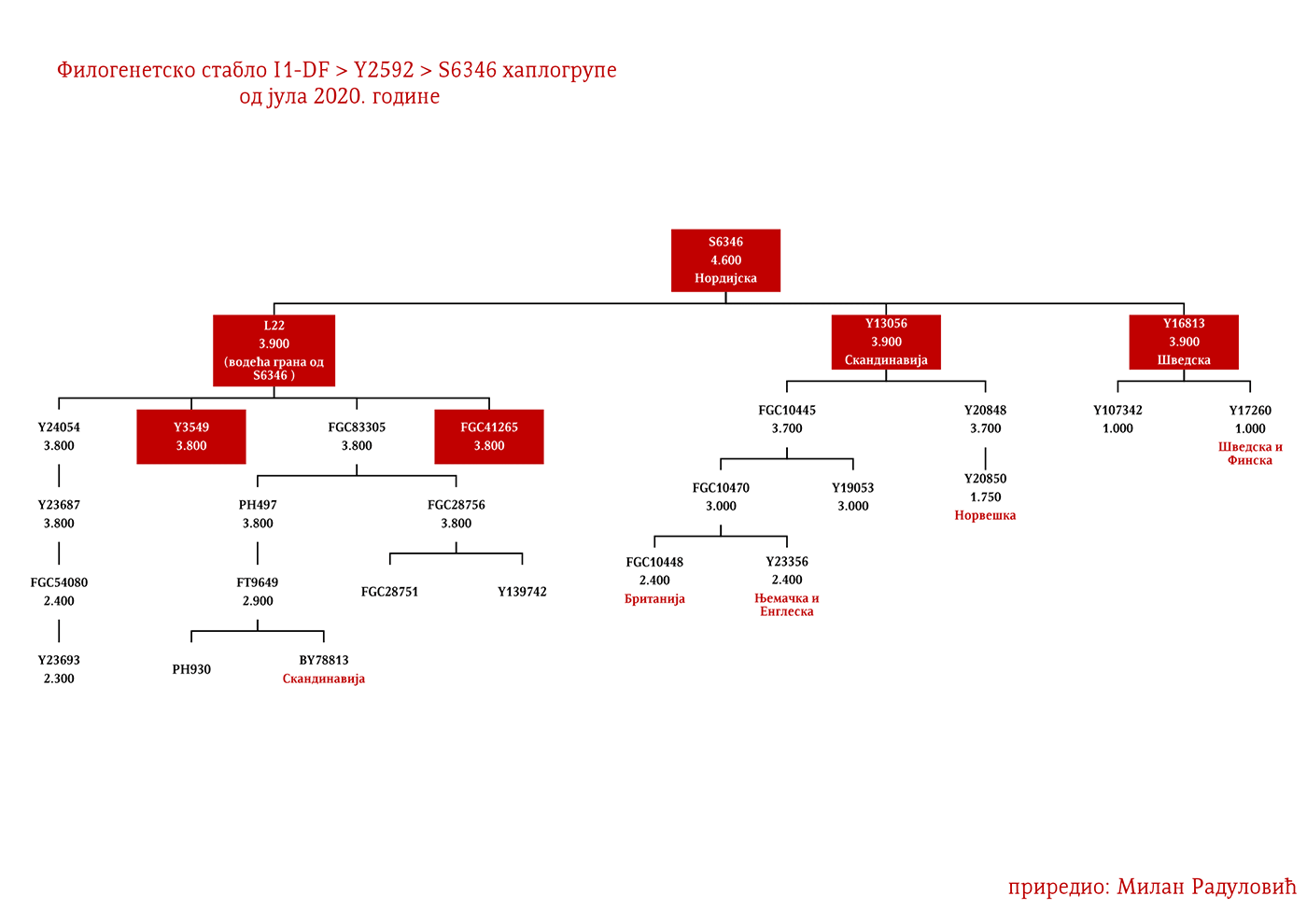
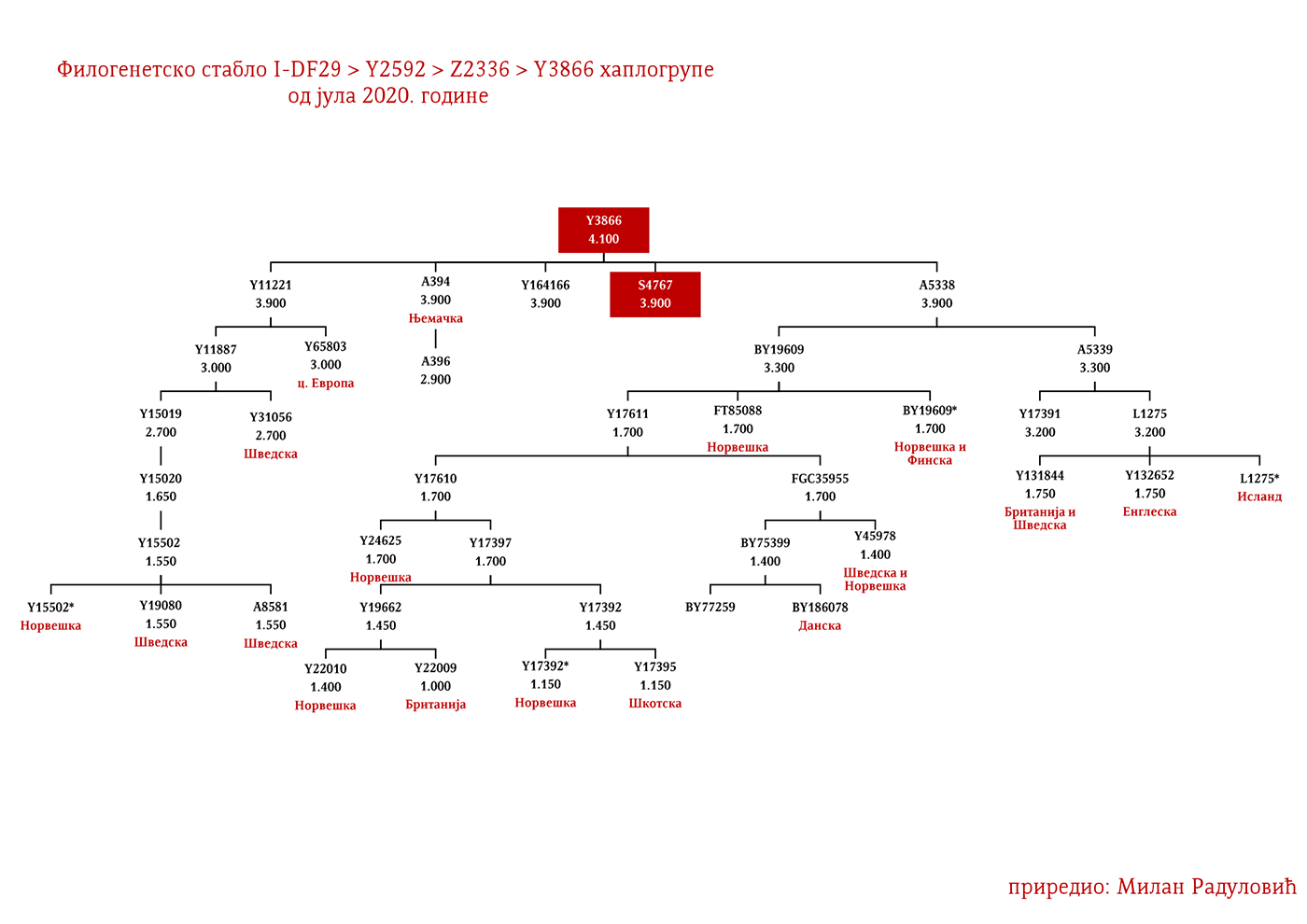
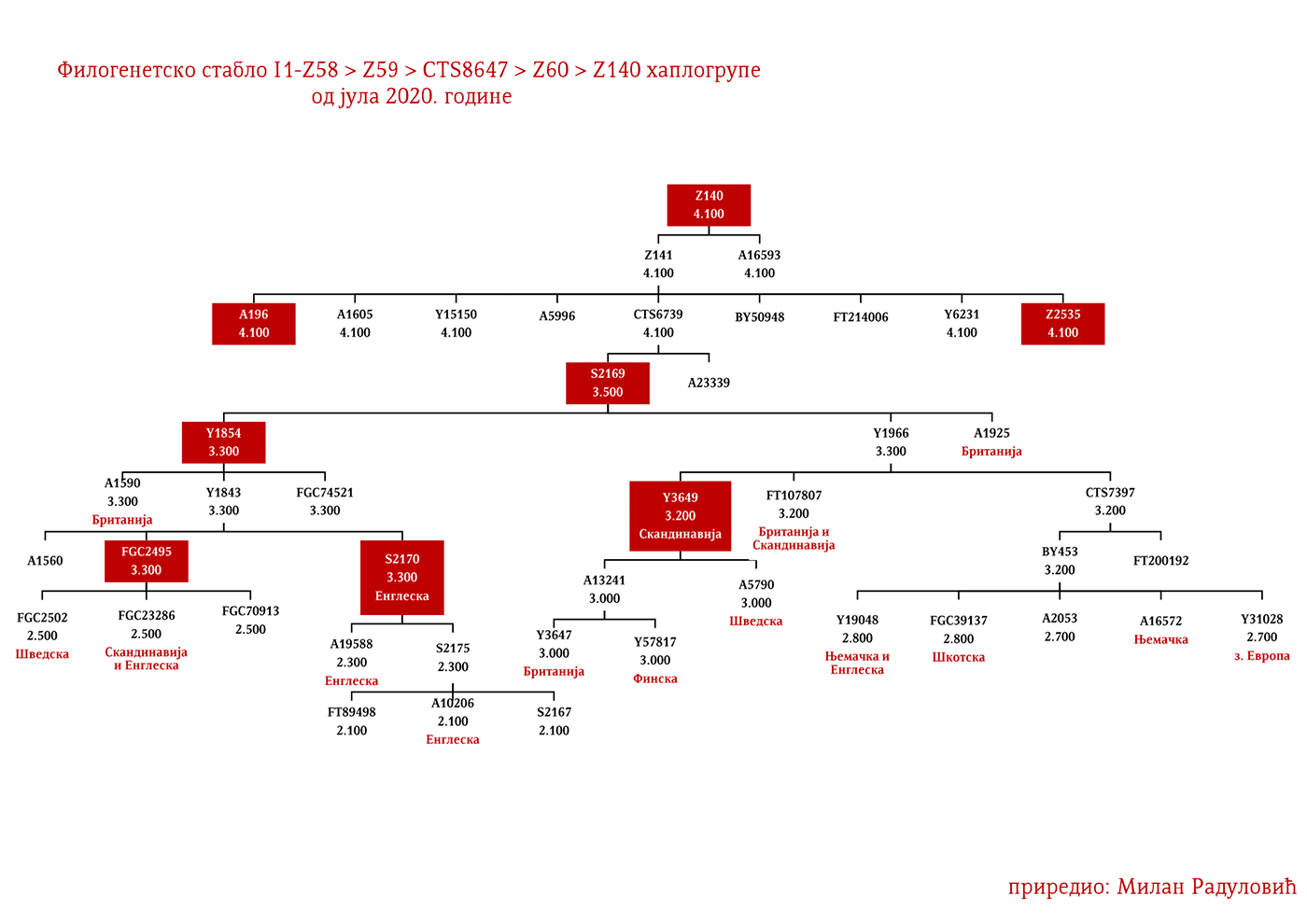
Dakle, od 37 uzoraka njih četiri je iz Švedske, a od toga dva uzorka pripadaju I1 haplogrupi, međutim oni nisu bili deo Kulture vrpčaste keramike već koegzistirajuće kasnoneolitske kulture. Imajući u vidu kasnije nalaze, nema sumnje da su se utopili u ovu kulturu i da su činili dobar dio njenog ogranka poznatog pod nazivom Kultura bojnih sjekira. Sa dolaskom pripadnika Kulture vrpčaste keramike u Skandinaviju, nastupio je eneolitski period bez neke posebne neolitske tranzicije i najavljeno je bronzano doba. Ujedno vrijedi pomenuti da je ovo vrijeme početka širenja I1 populacije, odnosno nastanka grana I1-CTS12768 (koja za sada nije uočena od strane ISOGG), I1-Z17954 i I1-DF29 koja se ubrzo razgranala na niz ogranaka. Sa područja sjevera centralne Evrope sledeće grane od I1-DF29>Z63 su započele širenje centralnom i zapadnom Evropom: I1-Z63>BY62 i BY151>FGC81364, kao i grane od I1-Z58, I1-Z58>A6867 i I1-Z58>PH4482, dok I1-L22 započinje naglo širenje među protonordijskom populacijom, a ostale grane I1 doživjeće širu ekspanziju centralnom Evropom mahom tokom perioda 1800-1200. godine p.n.e, sudeći po vremenu nastanka grana, njihovoj raznovrsnosti i zastupljenosti, te vremenu do zajedničkog pretka.
Već u narednom periodu posle 1700. godine p.n.e. uslediće novi talas Indoevropljana koji će Skandinaviji donijeti germanski karakter.
BRONZANO DOBA
Nordijsko bronzano doba počinje 1700. godine p.n.e. i završava se 500. godine p.n.e, a njega karakteriše novi talas doseljenika iz centralne Evrope oličenih u haplogrupama karakterističnijim za germanske narode. Tada započinje naglo širenje I1 haplogrupe na tlu centralne Evrope, sa intenziviranjem komunikacije sa kontinentalnim dijelom. Brodovi postaju glavna sredstva komunikacije i trgovine, njihov značaj vidi se pored ostalog i u tome što su Nordijci tokom bronzanog doba pravili spomenike u vidu „kamenih brodova“ (Kane, 2019) na grobnim humkama.
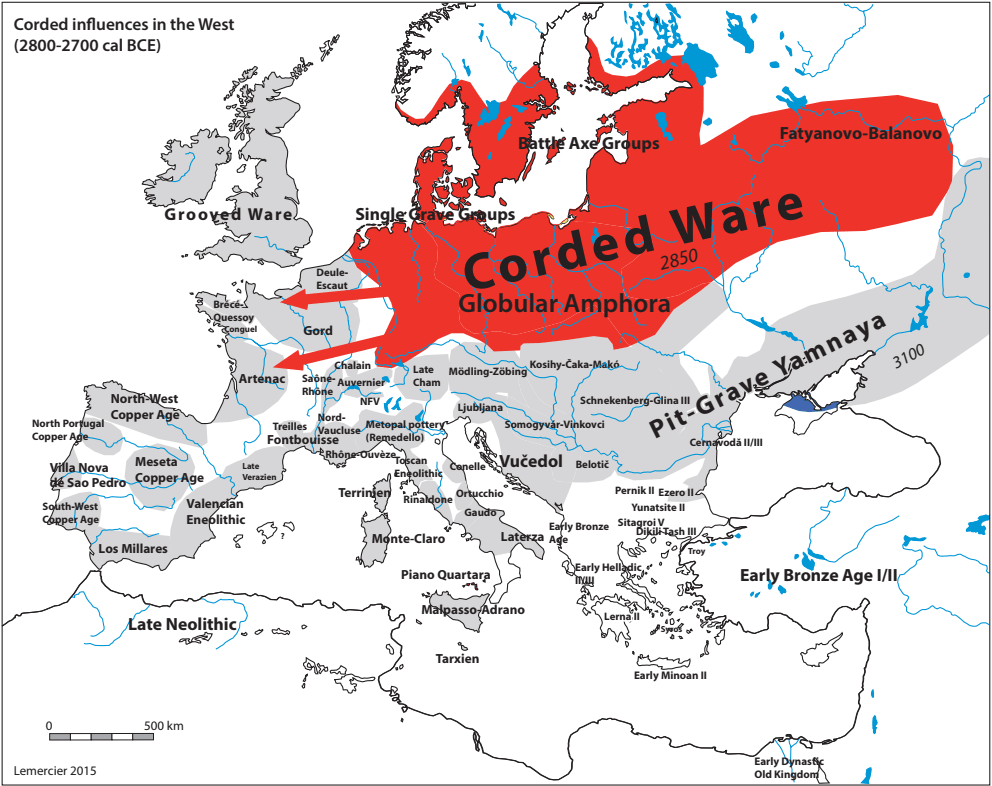
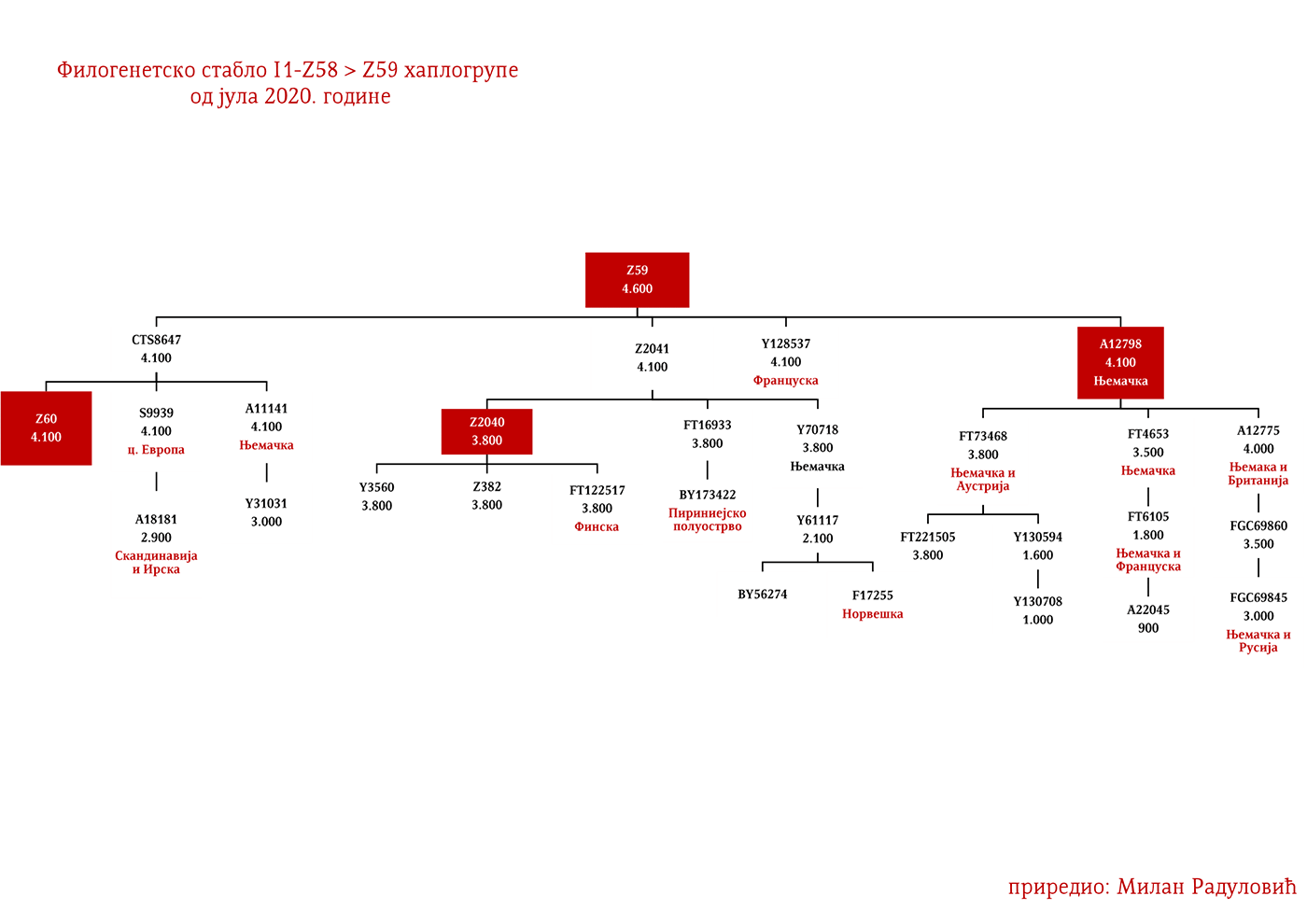
Drugi talas Indoevropljana vrlo je vjerovatno posledica ekspanzije Unjetičke kulture (2300-1600. god. p.n.e.) sa prostora današnje Njemačke. Odatle su se pripadnici R1b-U106 haplogrupe proširili na sjever, gdje su prvi datirani u 1400. god. p.n.e. prema rezultatima iz studije Alentoft i saradnika. Sama Unjetička kultura slabije je pokrivena genetičkim studijama, testirano je svega 14 (I5043, I0804, I7197, I0114, I0116, I7199, I4884, I5037, RISE145.SG, I5042, RISE431.SG, I7203, I7196, I7202, uzorci iz pomenutih studija za Unjetičku kulturu) njenih muških pripadnika. Uglavnom su je sudeći po uzorku (Olalde, et al., 2018; Allentoft, et al., 2015; Mathieson, et al., 2015) sačinjavali I2a2, R1a i R1b muškarci, sa manjim udjelom G2a haplogrupe. Tokom pomenutog perioda odigrala se prva etnogeneza germanskih naroda i u njoj su učestvovali pripadnici I1 haplogrupe. Ujedno ovo je, kako smo prethodno naveli, period naglog širenja njenih podgrana. U ovom periodu su sledeće podgrane izvršile uočljivije pozicioniranje: Z63 koja je mahom koncentrisana u centralnoj Evropi na prostoru današnje Njemačke, i izostaje u nordijskoj populaciji; Z58 koja je koncentrisana u zapadnim djelovima centralne Evrope; L22 koja je grupisana u Skandinaviji, kao i Z2336, dok je M227 smještena istočnije. Kako se vrijeme širenja podudara sa etnogenezom germanskih naroda, vrijedi pogledati kartu iz Priloga 5 na kojoj je prikazana ekspanzija Germana, koja je relevantna za I1 haplogrupu.
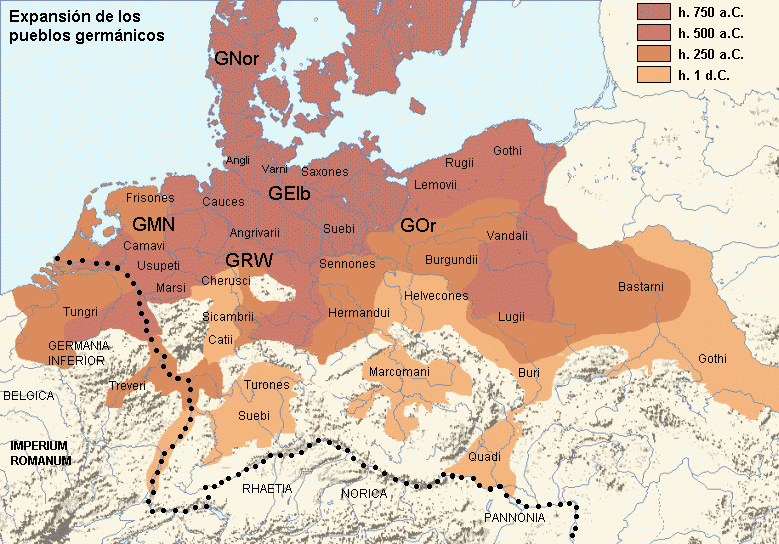
Od 6 uzoraka drevne DNK iz nordijskog bronzanog doba iz Švedske, 4 su I1 haplogrupe i to su uzorci RISE179, RISE207, RISE210, RISE175, dok su 2 R1b haplogrupe. Na migracije iz pravca juga Skandinavije ka centralnoj Evropi vjeruje se da je bitno uticala nagla klimatska promjena koja se desila u periodu između 850. i 650. godine prije nove ere, kada je nastupilo naglo zahlađenje (Kane, 2019).
GVOZDENO DOBA I ANTIKA
Kako je fokus haplogrupe na Skandinaviji, gvozdeno doba dijeli se na predrimsko gvozdeno doba koje počinje 500. godine p.n.e. i traje do 1. godine nove ere, drugi period je rimsko gvozdeno doba koje traje od 1. do 400. godine n.e, zatim sledi germansko gvozdeno doba od 400. do 800. godine.
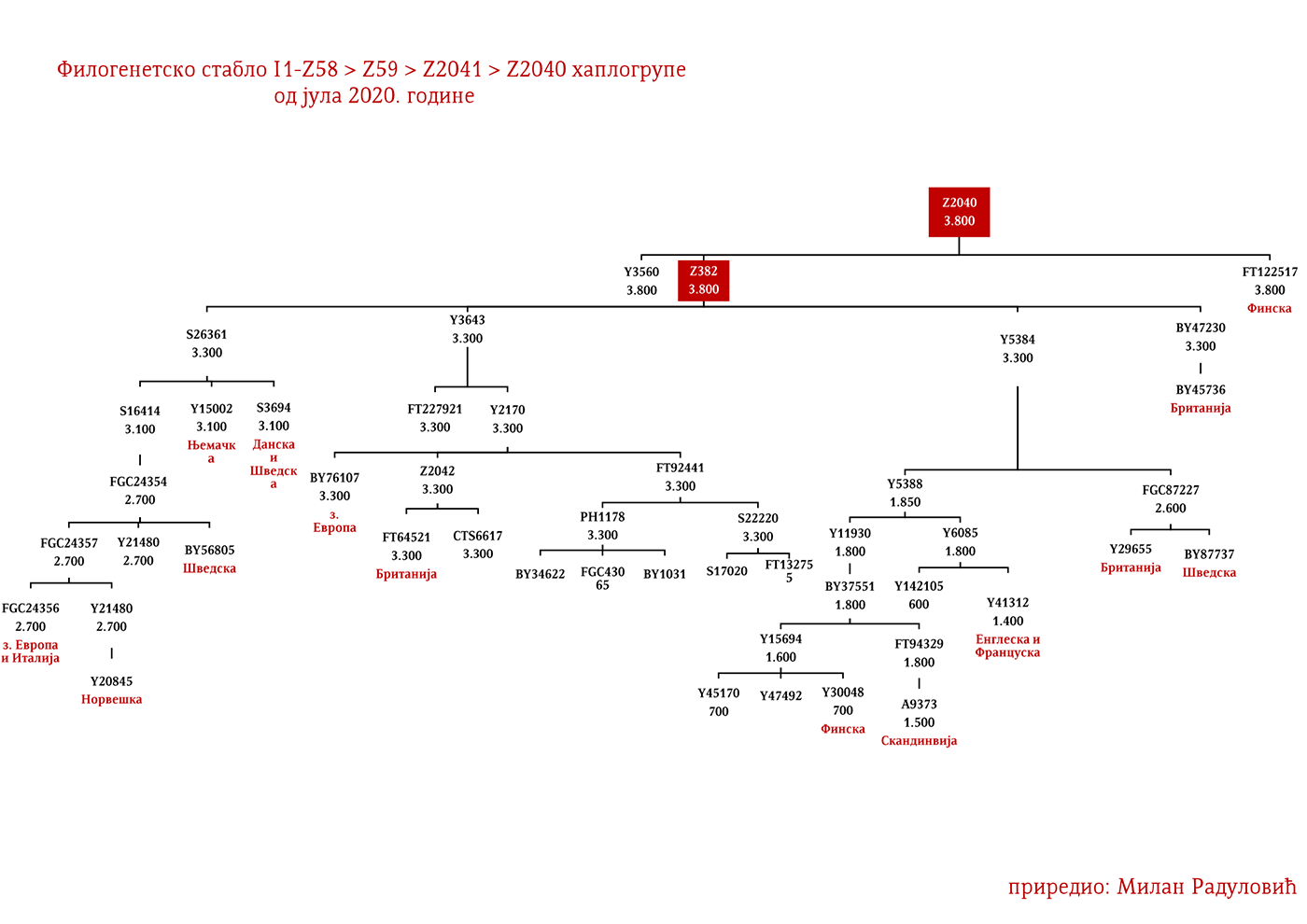
Najstariji uzorak kojim raspolažemo iz gvozdenog doba je sa juga Francuske iz Besana, a pripadao je latenskoj kulturi (450-1. god. p.n.e.). Nalazi se pod šifrom BES1248 i datiran je između 200-50. god. p.n.e. (Brunel, et al., 2020), zatim VK521 iz Danske (Margaryan A. , et al., 2019), pripadnik I1-Z58>Z2040 haplogrupe koji je datiran u 300. godinu. U centralnoj Evropi iz ovog perioda evidentirani su uzorci AED_249, STR_241 i STR_486 iz rada Veramaha i saradnika (Veeramah, et al., 2018), koji potiču iz Bavarske i datirani su u period između 460-530. godine. Kod ovih uzoraka karakteristično je to što su svi imali potpune sjevernocentralne evropske genetičke primjese (Fig. 3. iz studije). Uzorak AED_249 pripadao je podgrani I1-DF29 (xP109), STR_241 je I1-DF29 (xM227, Z138, Z60) dok je STR_486 I1-M253 (xP109). Zatim imamo uzorak KO_55 iz Poljske koji pripada vilbarskoj kulturi (Zenczak, et al., 2017), datiran u 100-300. godinu, koji pripada podgrani I1-Z63>L1237 (Rajevac, 2020). U Mađarskoj na lokalitetu Solad (Szólád), na uzorku od 21 skeleta jedan je I1-P109>FGC21810 (šifra uzorka je SZ45), dok su ostali dominantno R1b i I2a2 (Amorim, et al., 2018) muškarci. Na prostoru Mađarske nađen je još jedan I1-M253 uzorak ali iz ranog avarskog perioda, SzO/540 (Neparáczki, et al., 2019). Priloženi uzorci iz radova dovode do zaključka o raznovrsnosti I1 podgrana u centralnoj Evropi koja odgovara njihovoj današnjoj rasprostranjenosti, što se podudara sa ranije napisanim zaključcima u vezi ove haplogrupe. Uzorci Langobarda iz kasnoantičkog perioda iz Italije, R110 koji pripada podgrani I1-Z63>Y7234 i CL63 koji pripada podgrani Y2245, zajedno sa BES1248 sa juga Francuske, svjedoče o migracijama germanskih plemena. Anglosaksonski uzorak NO3423 sa područja Velike Britanije, koji pripada podgrani I1a-DF29 (xZ63, M227,Y3603, Z2041, Z138, CTS7362, Z141), s druge strane svjedoči o zapadnim dosezanjima I1 muškaraca tokom kasnoantičkog perioda.
SNP MUTACIJE
Haplogrupu I1 definišu snipovi M253, P30, P40, M307 i M450 (Karafet, et al., 2008). Ona se prema trenutnom ISOGG stablu 2019 dijeli na tri grane: I1a koju definiše snip DF29, I1b definisana Z131 i I1c definisana Z17943 snipom (ISOGG, 2020). Ovde ćemo konstatovati da ISOGG kasni sa osvježavanjem stabla, jer Z17943 i Z131 imaju zajedničkog pretka Z17954, a koji je na istom filogenetskom nivou sa DF29 i novotkrivenom linijom CTS12768, stoga bi u budućoj osvježenoj verziji stabla Z17954 trebala nositi oznaku I1b dok bi CTS12768 nosio oznaku I1c.
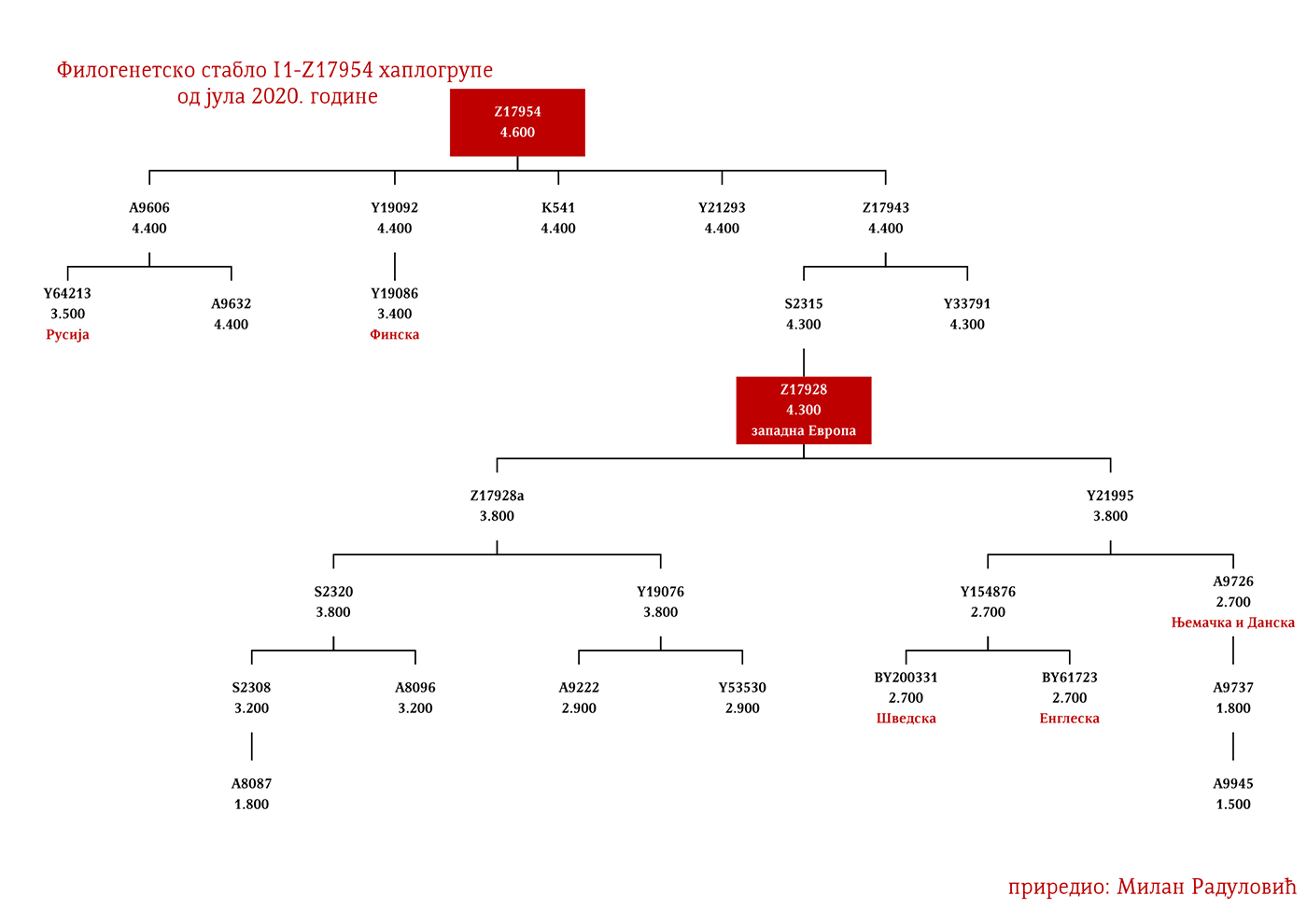
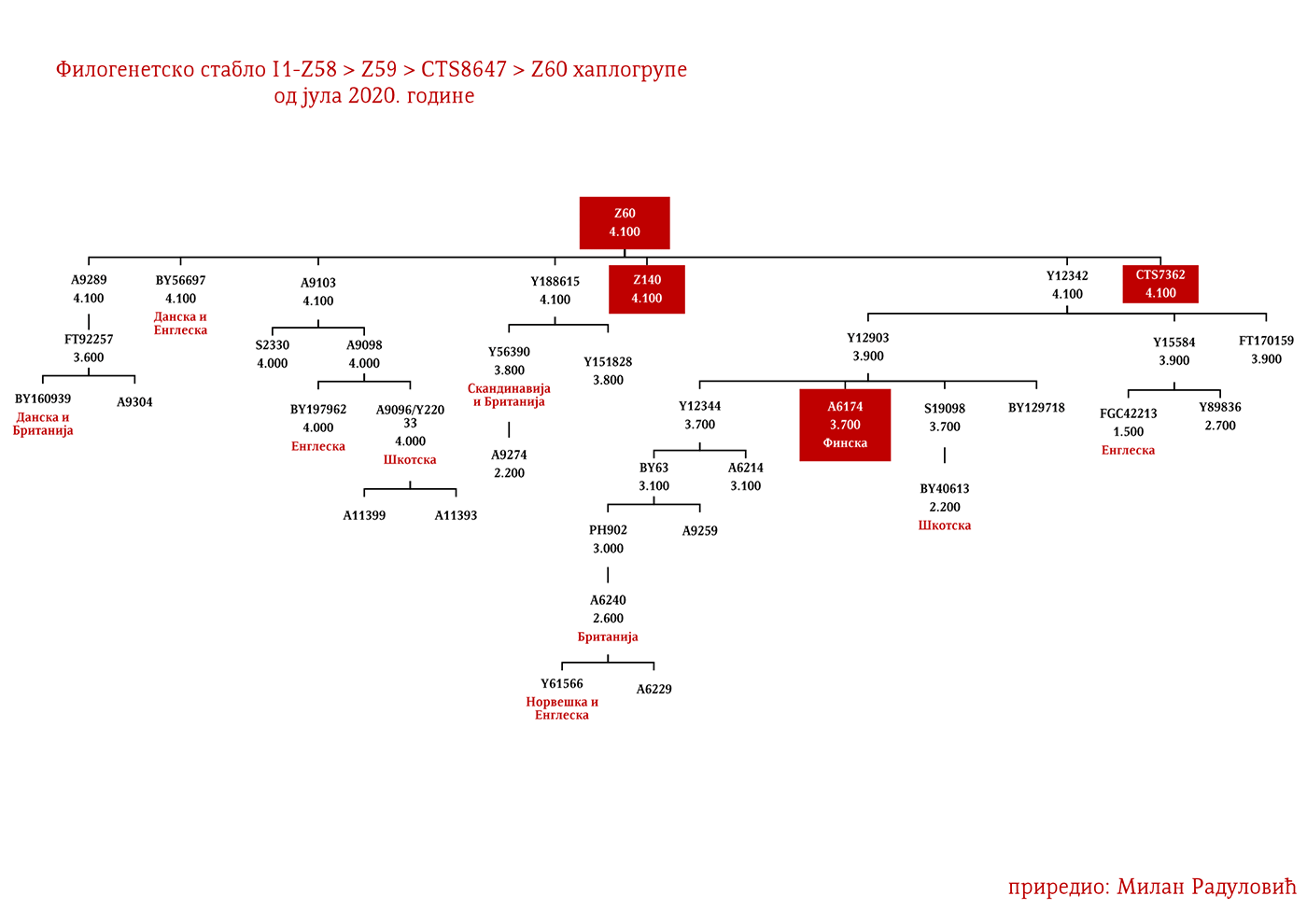
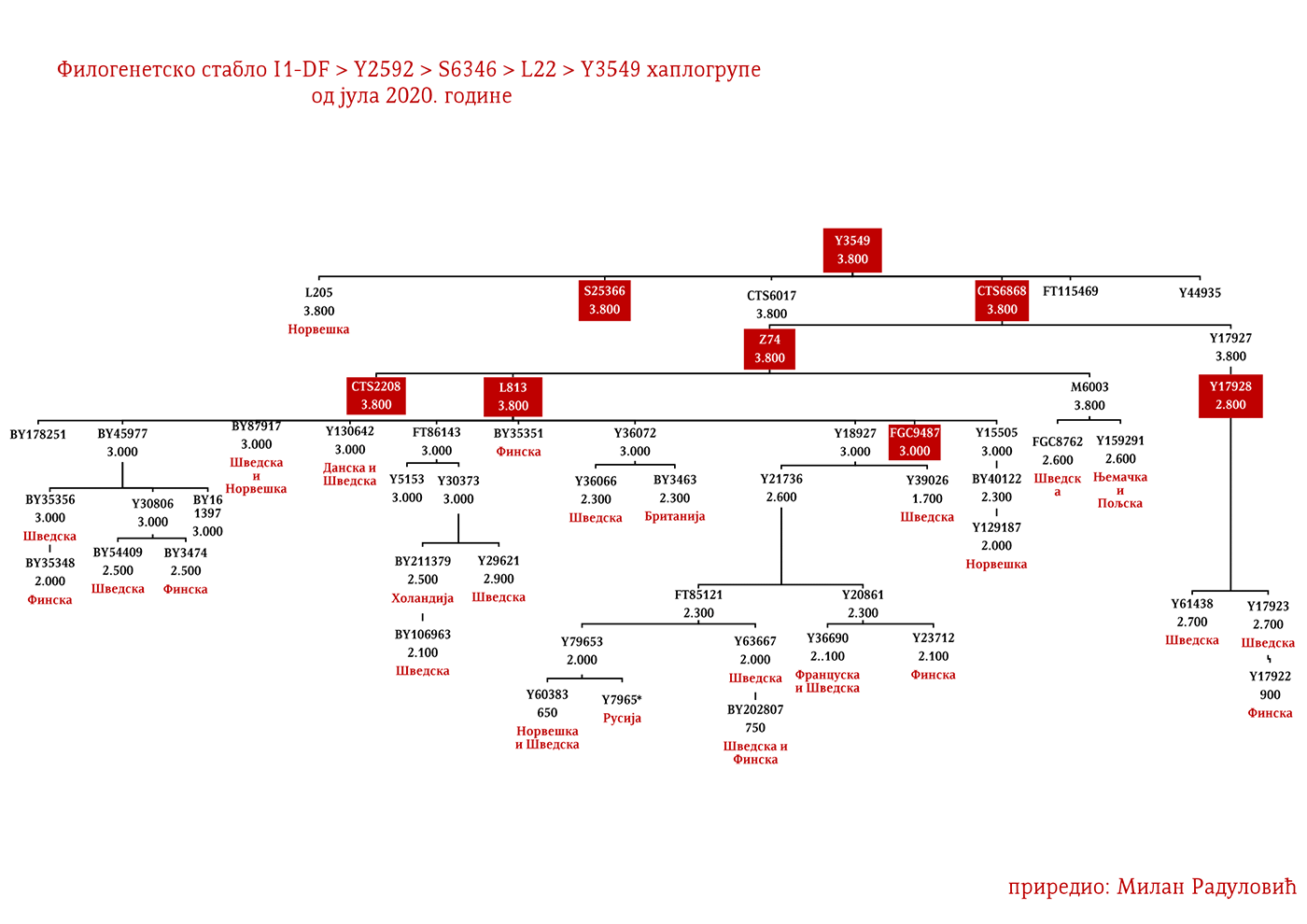
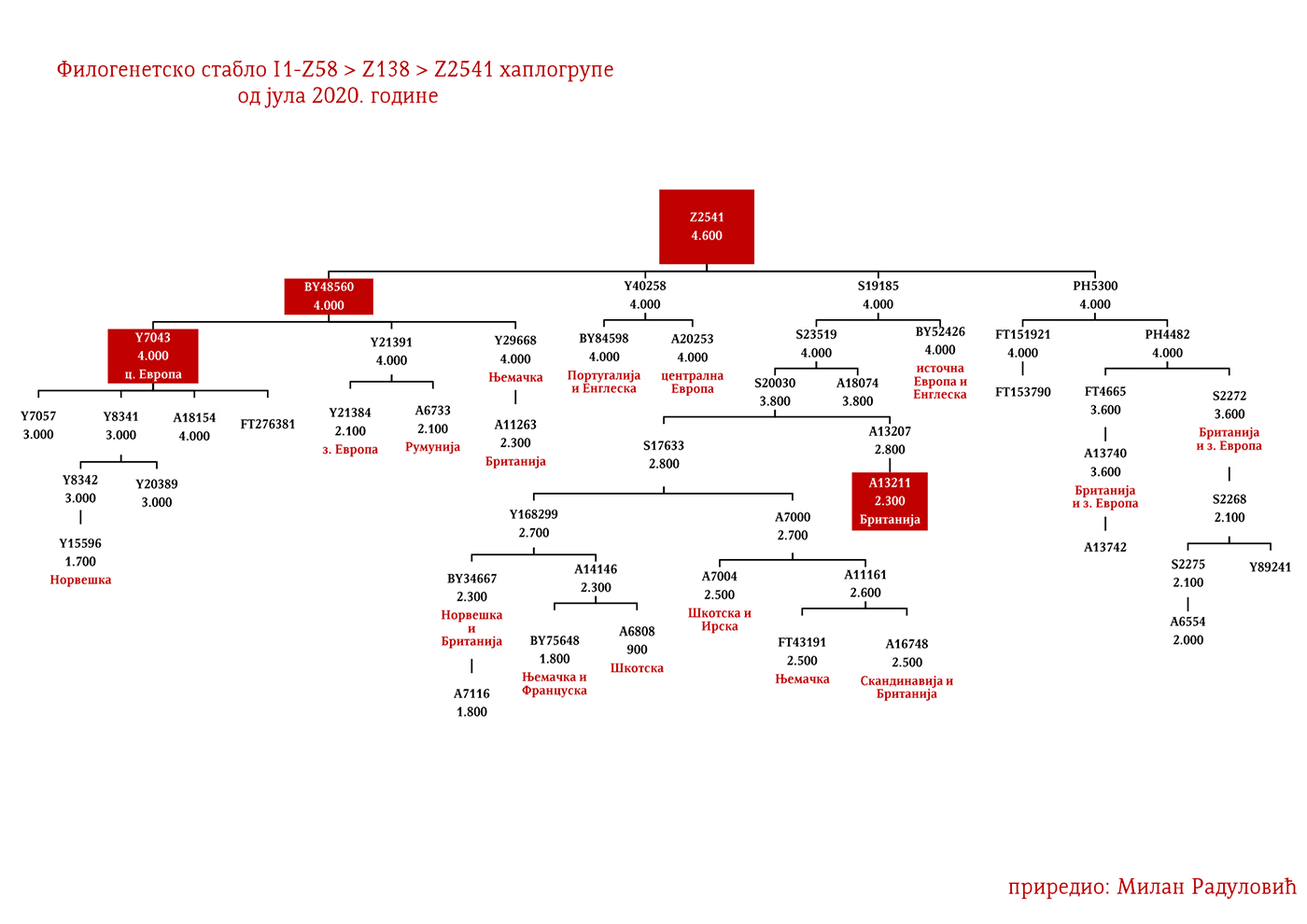
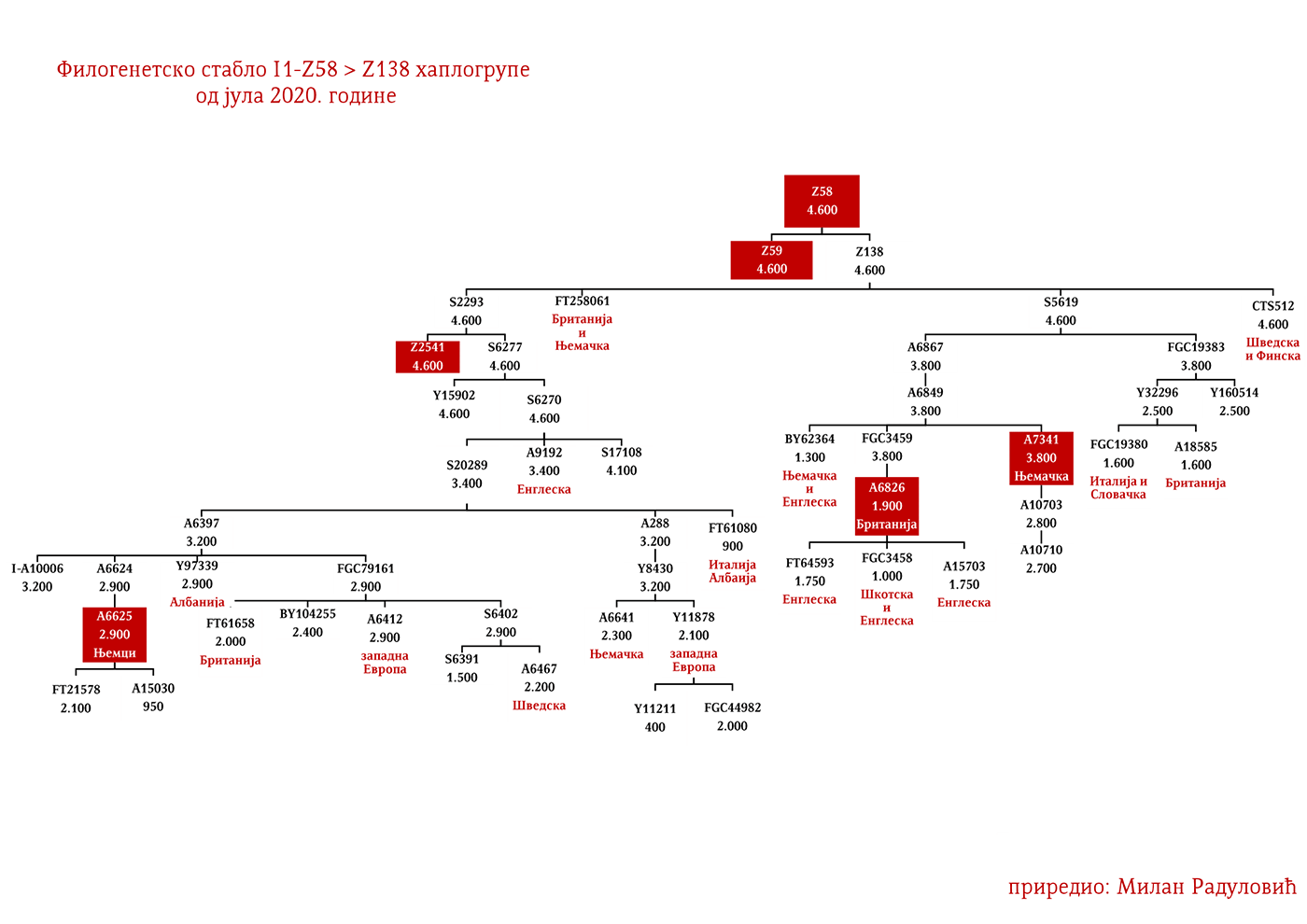
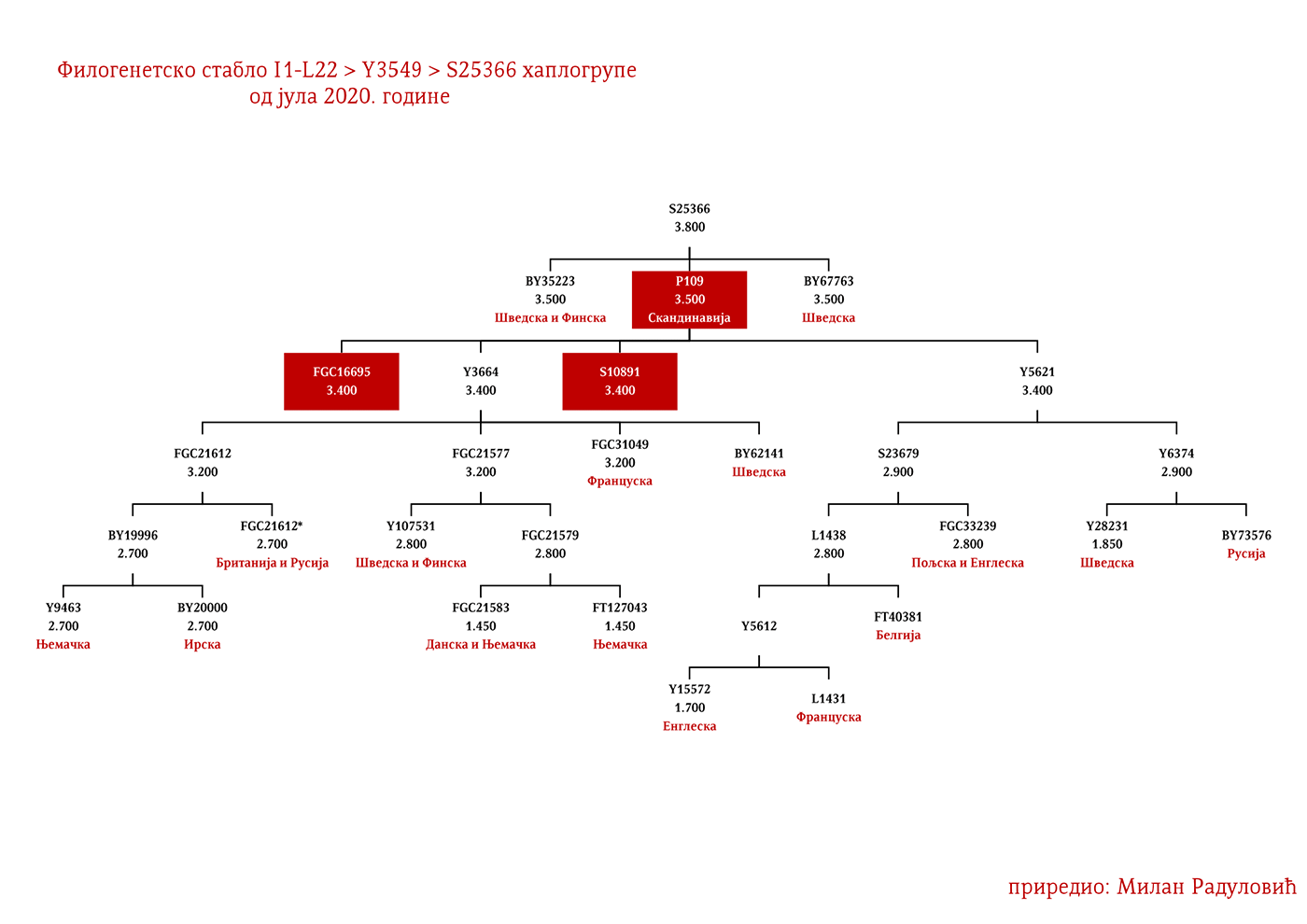
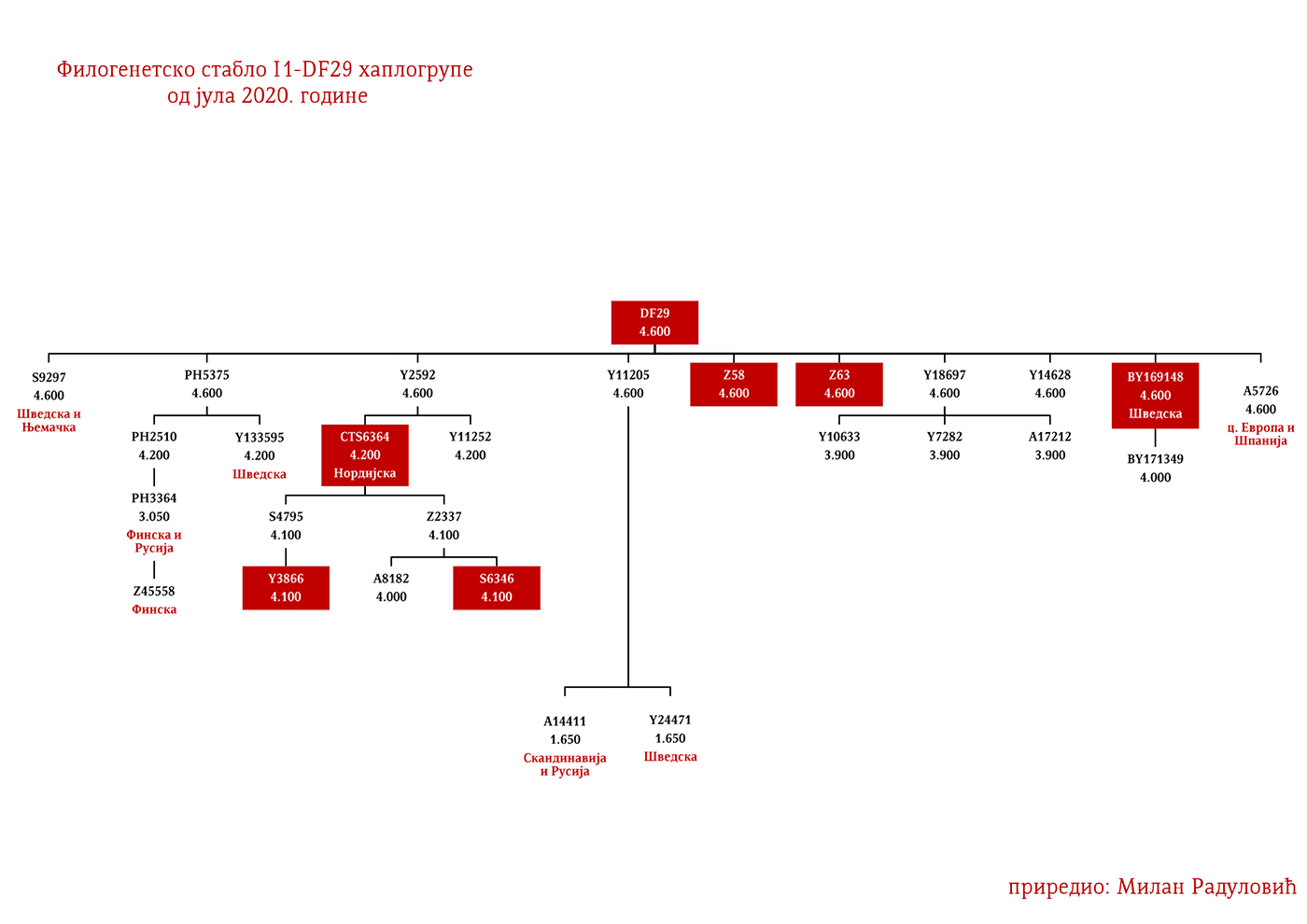
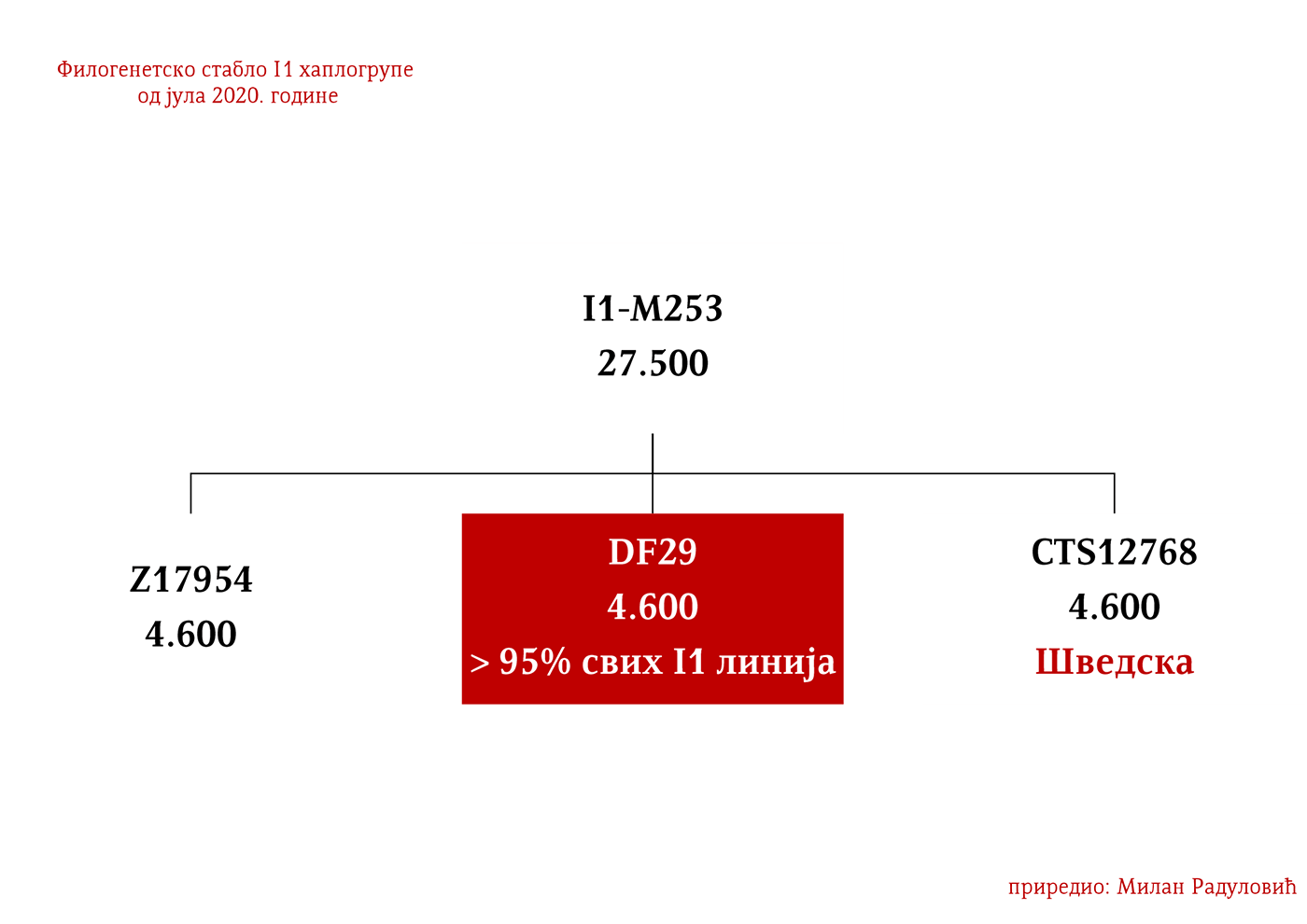
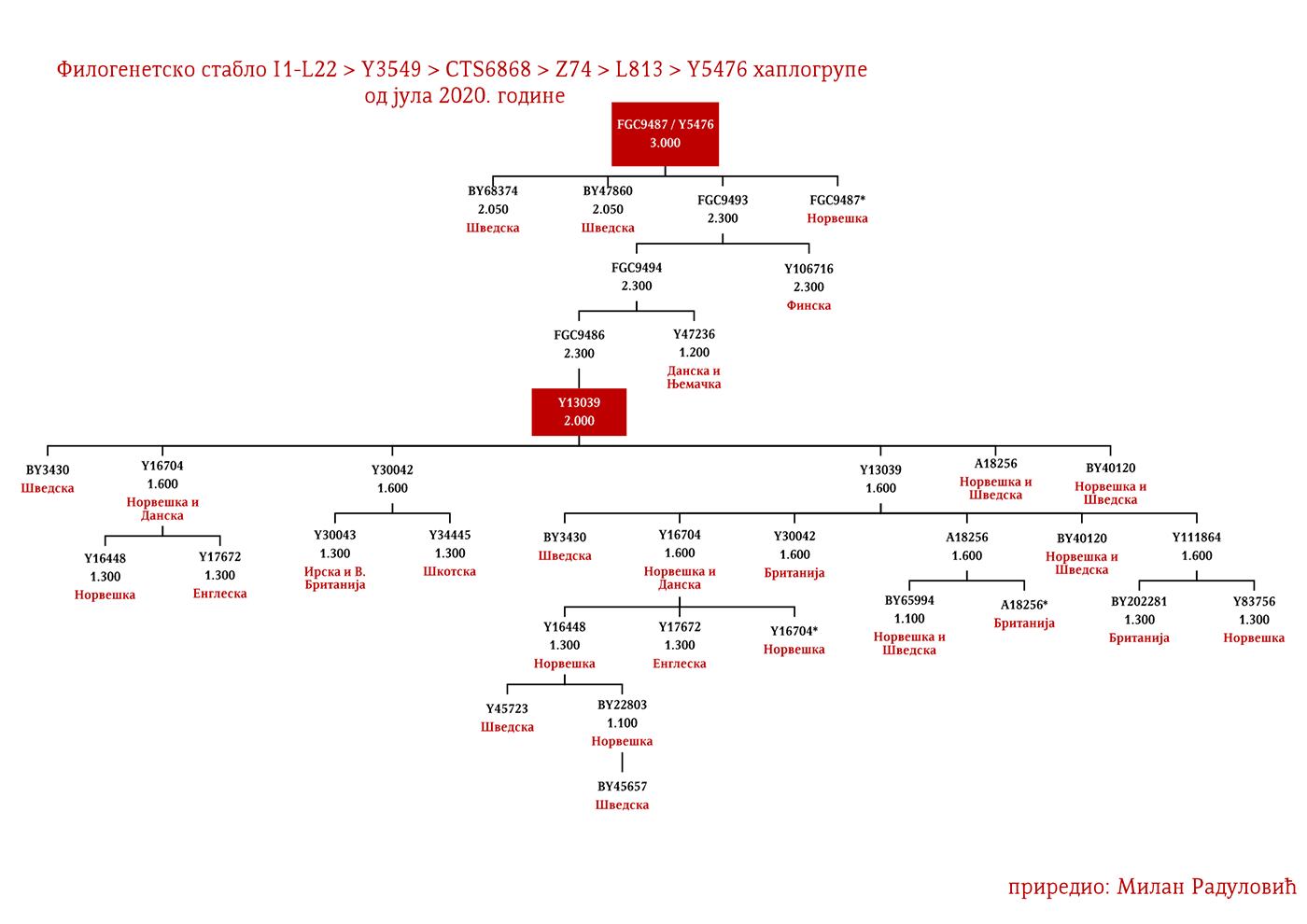
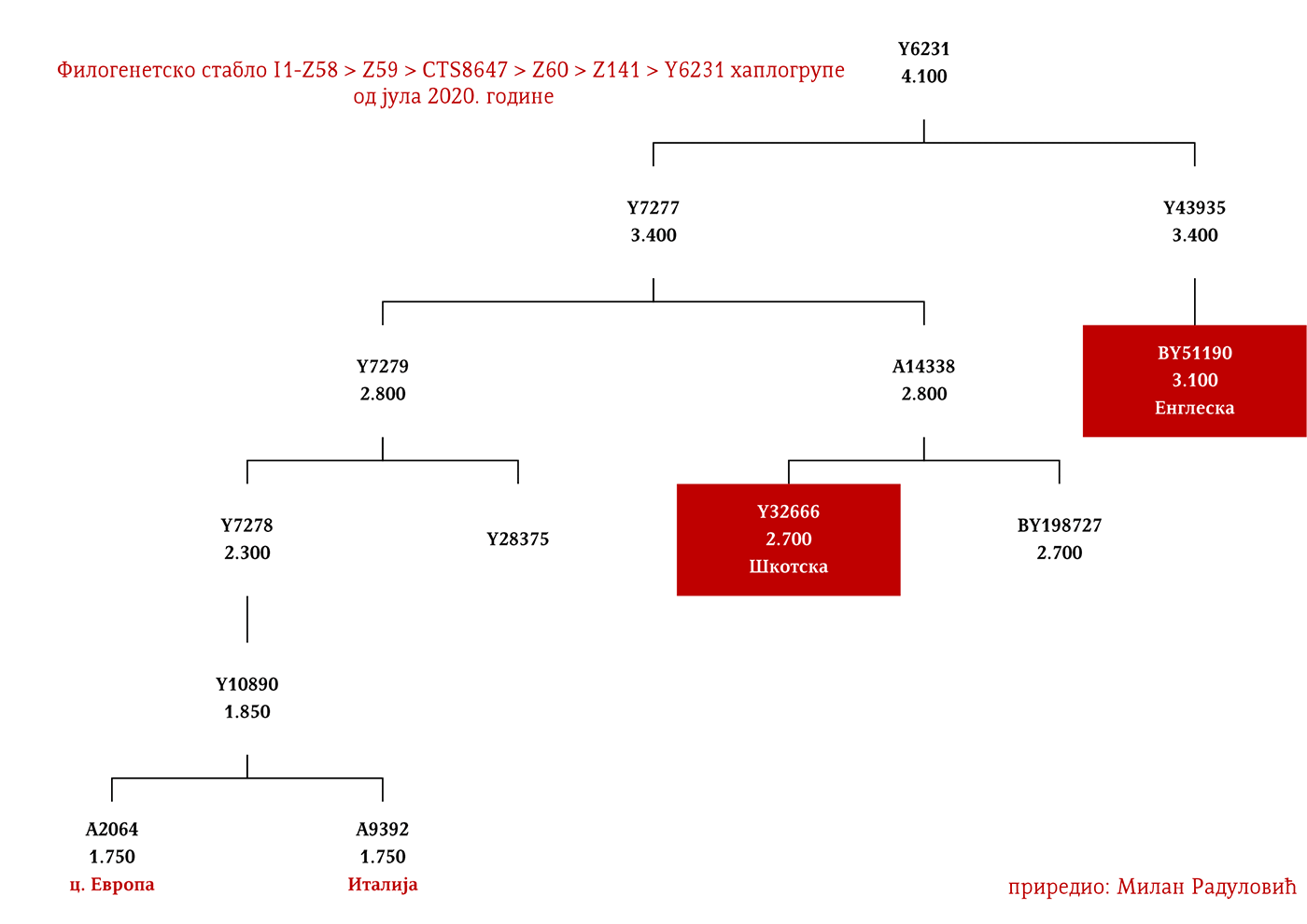
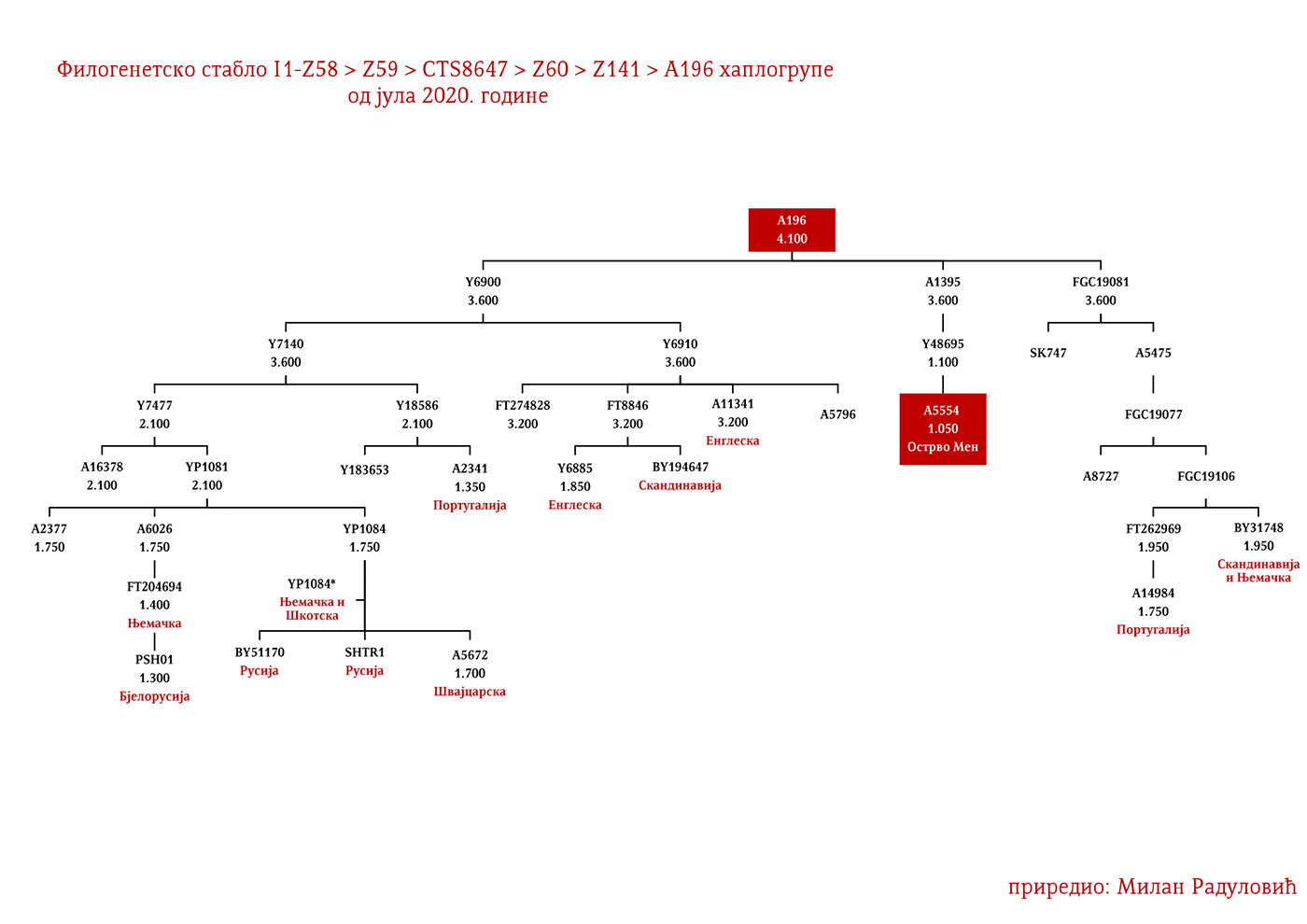
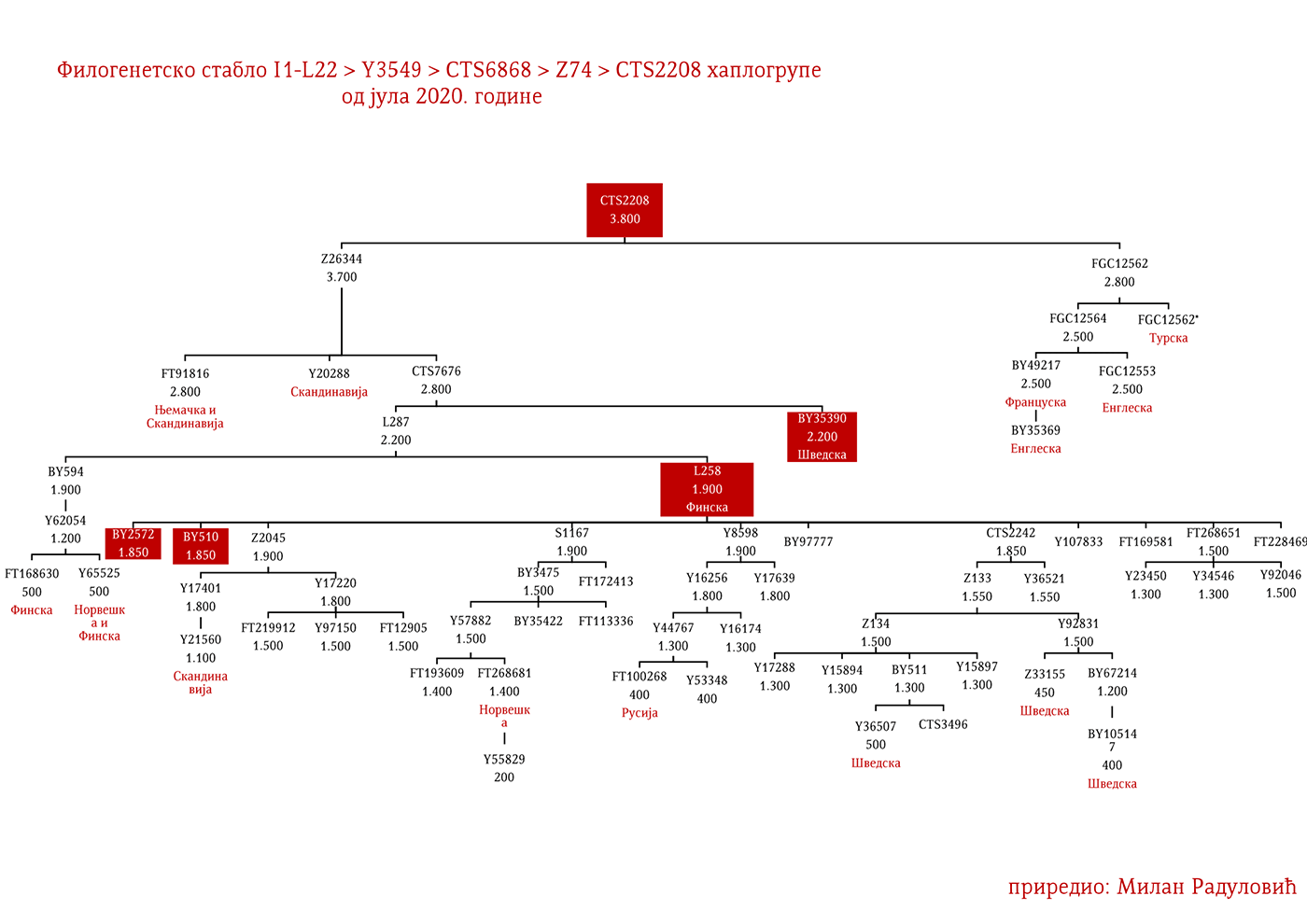
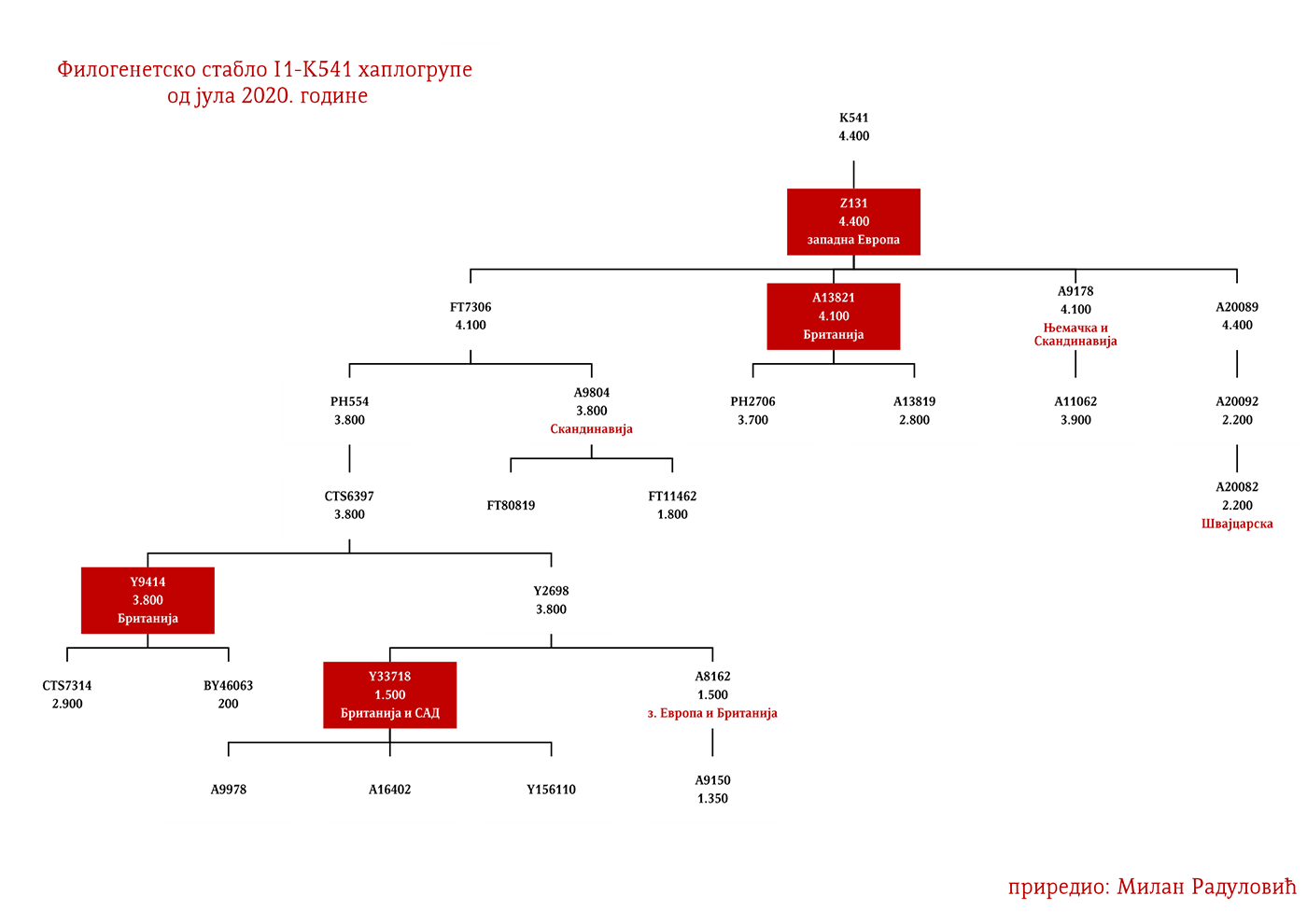
ZASTUPLJENOST I PODGRUPE
Haplogrupa I1 zastupljena je u Evropi gdje na sjeveru kontinenta bilježi najveću zastupljenost i raznovrsnost grana haplogrupe. Procentualno gledano najzastupljenija je u nordijskim i germanskim zemljama. U Norveškoj je zastupljena sa 39%, Švedskoj sa 35%, Islandu 34%, Danskoj 32%, Finskoj 23%, Njemačkoj 17%, Holandiji 16%, Austriji, Belgiji, Estoniji 12%, Velikoj Britaniji 11%, dok je kod srpske populacije zastupljena sa 8% (prema podacima Srpskog DNK projekta).
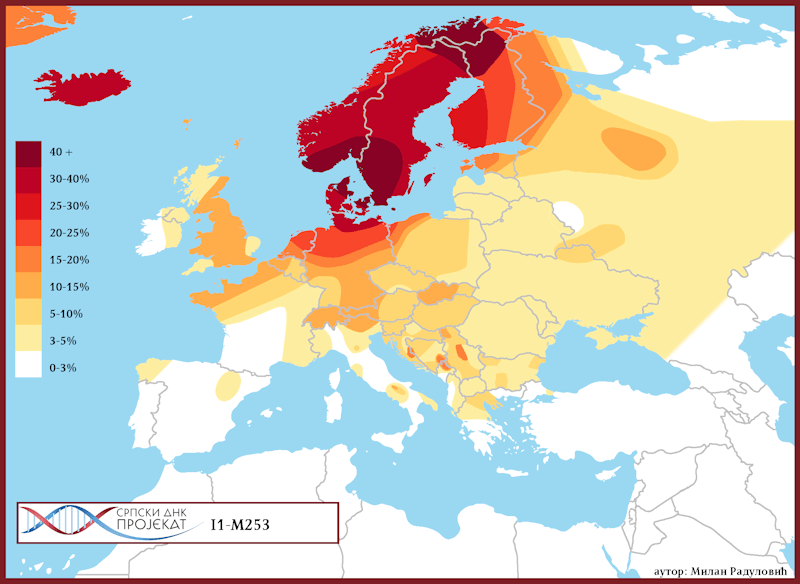
Detaljan pregled zastupljenosti haplogrupe I1 u evropskim populacijama možemo videti u Prilogu 7 ispod.
U .pdf verziji dat je detaljan prikaz grana haplogrupe, kao i njihovo geografsko i etničko gravitiranje, pri čemu još jednom skrećemo pažnju da je većina podgrana zastupljena i nordijskim zemaljama i kod germanskih naroda.
STARI NARODI
Haplogrupu I1 možemo povezati sa germanskim narodima, ona je učestvovala u njihovoj etnogenezi zajedno sa R1b-U106, R1b-L238, I2a2-M223, R1a-Z284 i R1a-L664 (hYP942) haplogrupom, što je svojevremeno i potvrđeno arheogenetikom.
VIKINZI
Predstavljaju grupu naroda sa sjevera Evrope iz zemalja Danske, Norveške i Švedske koji su najprije započeli ratničke pohode ka zapadnoj Evropi, a nešto kasnije ka istoku prema slovenskom svijetu (gdje se tradicionalno nazivaju Varjazima). Vikinško doba, kako to nazivaju zapadnoevropski istoričari, počinje 793. godine sa napadom Vikinga na manastir Lindisfarn na sjeveru Engleske, a traje do 1066. kada su Normani u bitci kod Hejstingsa porazili engleskog kralja Harolda Godvinsona i postali vladajuća elita. U nedavno objavljenom radu o Vikinzima, na uzorku od 296 muških skeleta (Margaryan A. , et al., 2020), 92 su pripadala I1 haplogrupi, što je trećina od ukupnog uzorka. Od tog broja (92), 37 skeleta, odnosno 40%, pripadalo je I1-L22 linijama, čime je potvrđeno pisanje ranijih istraživača, Jerkovića (Jerković, Y-DNK haplogrupa I – najčešća kod Srba, 2012), Nordveta (podjela na „Norse” i „Ultra-Norse” haplotipove L22+) i Heja (Hay, Origins and history of Haplogroup I1 (Y-DNA), 2016) da je L22 najtipičnija vikinška haplogrupa. Pored L22+ linija, druga po učestalosti je Z59+ linija koja bi inače pretežno bila dobar kandidat za zapadnogermanske narode, Anglosaksonce. Upadljivo je odsustvo Z63 linija, što i ne čudi s obzirom da imaju gotovo isključivo kontinentalni karakter. Pronađena su i dva M227 uzorka, očekivano na istoku Švedske shodno osnovanoj pretpostavci da su uglavnom učestvovali u gotskim migracijama.
Moguće varjaške linije na osnovu rasprostranjenosti, arheogenetike (u tekstu kao VK), ishodišta i starosti grana, bile bi sledeće: I1-L22>Y21972, zastupljena u Rusiji i Švedskoj; I1-L22>Y31081, zastupljena u Ukrajini, Rusiji i Švedskoj; I1-L22>CTS2208*, Rusija, VK220; I1-L22>L813>Y79653, Rusija, Švedska i Norveška; I1-L22>CTS2208>FGC12562, Turska; I1-L22>Z26344>FT100268, Rusija i Finska; I1-L22>L813>FGC17580, nordijske zemlje i Rusija; I1-L22>P109>FGC21612, nordijske zemlje, Rusija i Engleska; I1-L22>CTS5350>BY65672, Y22486, Skandinavija i Rusija; I1-L22>Z2338, I1-L22>Y21959, I1-L22>CTS2208, I1-L22>L813>Y19207, I1-L22>Y5473, I1-L22>Y22024, I1-Z58>BY61100 – redom VK223, VK20, VK220, VK14, VK221, VK219, VK539.
GOTI
Na osnovu istih gore pomenutih parametara Goti bi bili I1-M227 i sledeće I1 linije: I1-Z58>S6402; I1-Z58>Y21391 dio; I1-Z58>A6867; I1-Z58>Z60>Z140>S4700; I1-Z63>S2078>L1242 dio; I1-Z63>S2078>Y16435; I1-Y2592>Y65803; I1-Z63>S2078>Y78357. Naravno ovo nije konačna ocjena, bilo je i drugih grana no za ove sa većom sigurnošću pretpostavljamo.
VIZIGOTI
Vizigotske linije bi mogle biti zaključujući prema gore pomenutom metodu: I1-Z63>S2078; I1-Z63>S2078>Y7666; I1-Z63>S2078>Y18660; I1-Z63>S2078>Y59728; I1-Y2592>Y164166; I1-Z58>Z60>Z140>Y29667; I1-Z58>Y7043.
DRUGI STARI NARODI
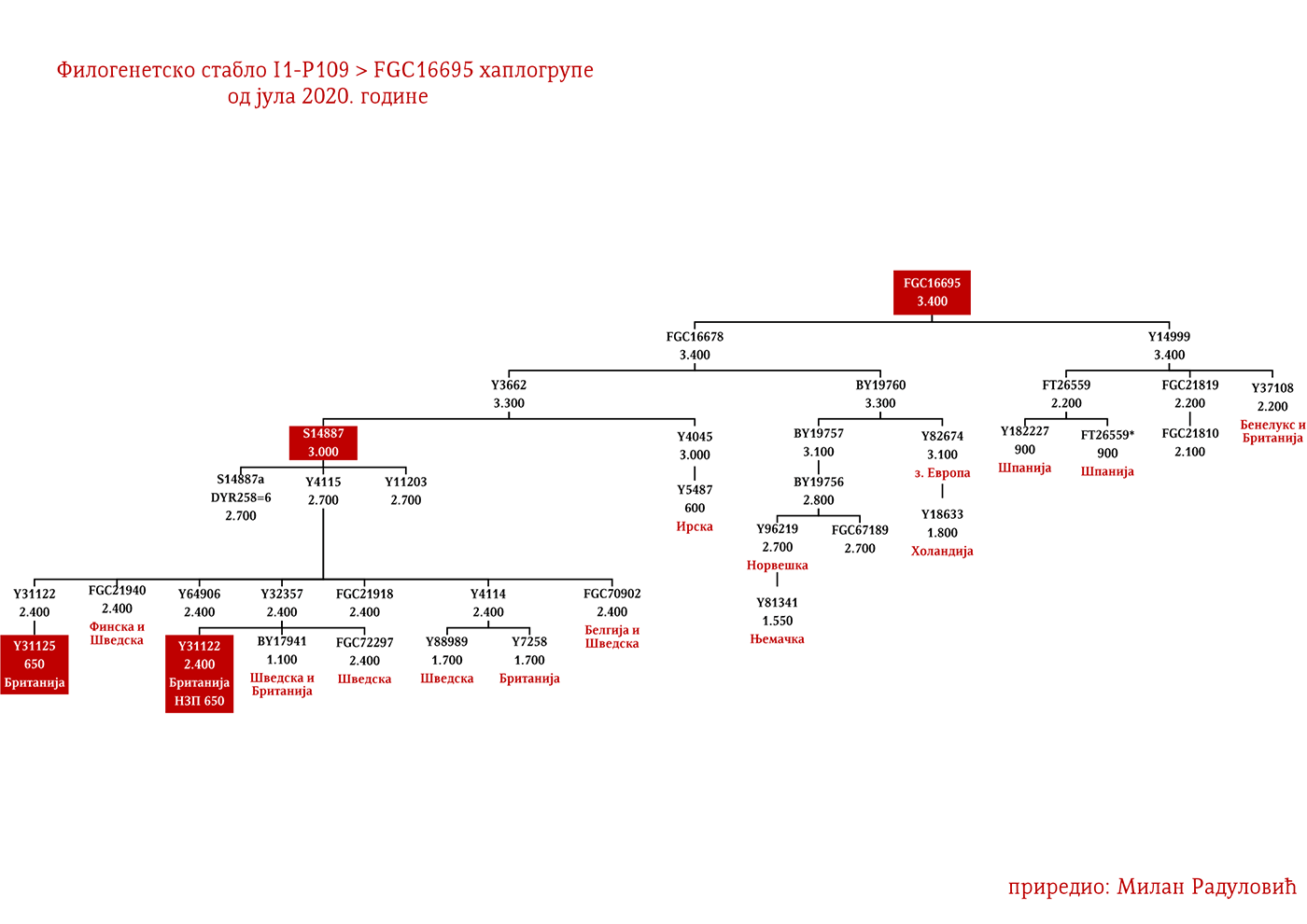
Zapadni germanski narodi bi mahom pripadali Z58 i Z63 linijama. Anglosaksonske i frizijske grane treba tražiti u onim podgranama koji se sreću u Engleskoj i Beneluksu, a koje imaju veću starost od 2000 godina. Za Langobarde imamo arheogenetsku potvrdu da je jedna njihova linija bila i I1-P109>FGC21810, uzorak SZ45 (Amorim, et al., 2018; Rajevac, 2020).
SRPSKI NAROD
Kod Srba najbrojni genetički I1 rodovi su Drobnjaci I1-P109>FGC22045 i Macure I1-Z63>Y16434, a slijede ih I1-Z63>Y13946>PH220 koji su rasuti na širokom području izemeđu Krajine na zapadu i zapadne Srbije na istoku; I1-P109>Y13930 se prostiru u dinarskoj zoni, a ishode iz istočne Hercegovine; I1-M227>A11380>BY169301 rasuti su od Krajine do Šumadije, a dovode se u vezu sa Gotima. Prisutne su naravno i druge grane, no ovo bi bile brojnije i genetički određenije.
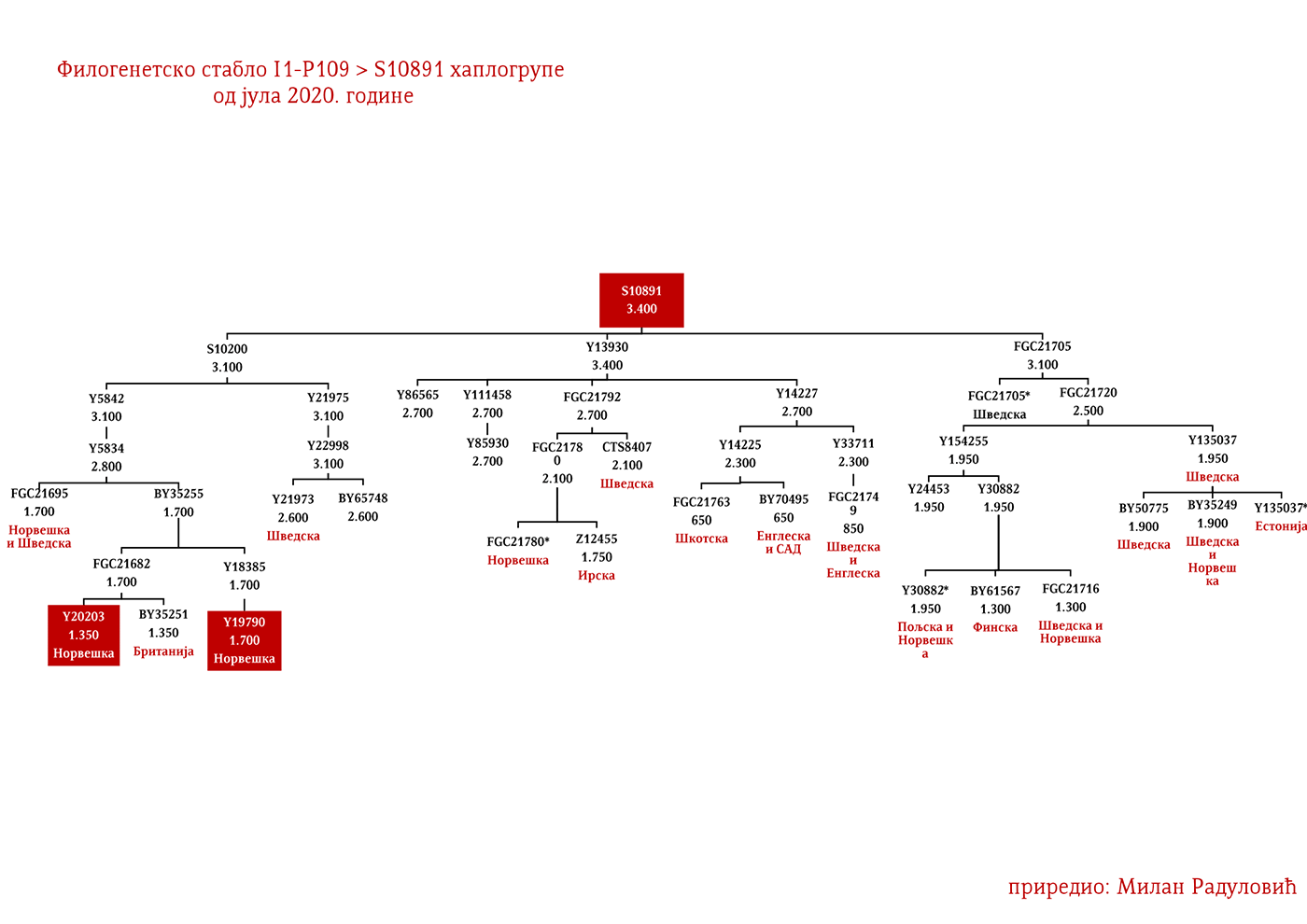
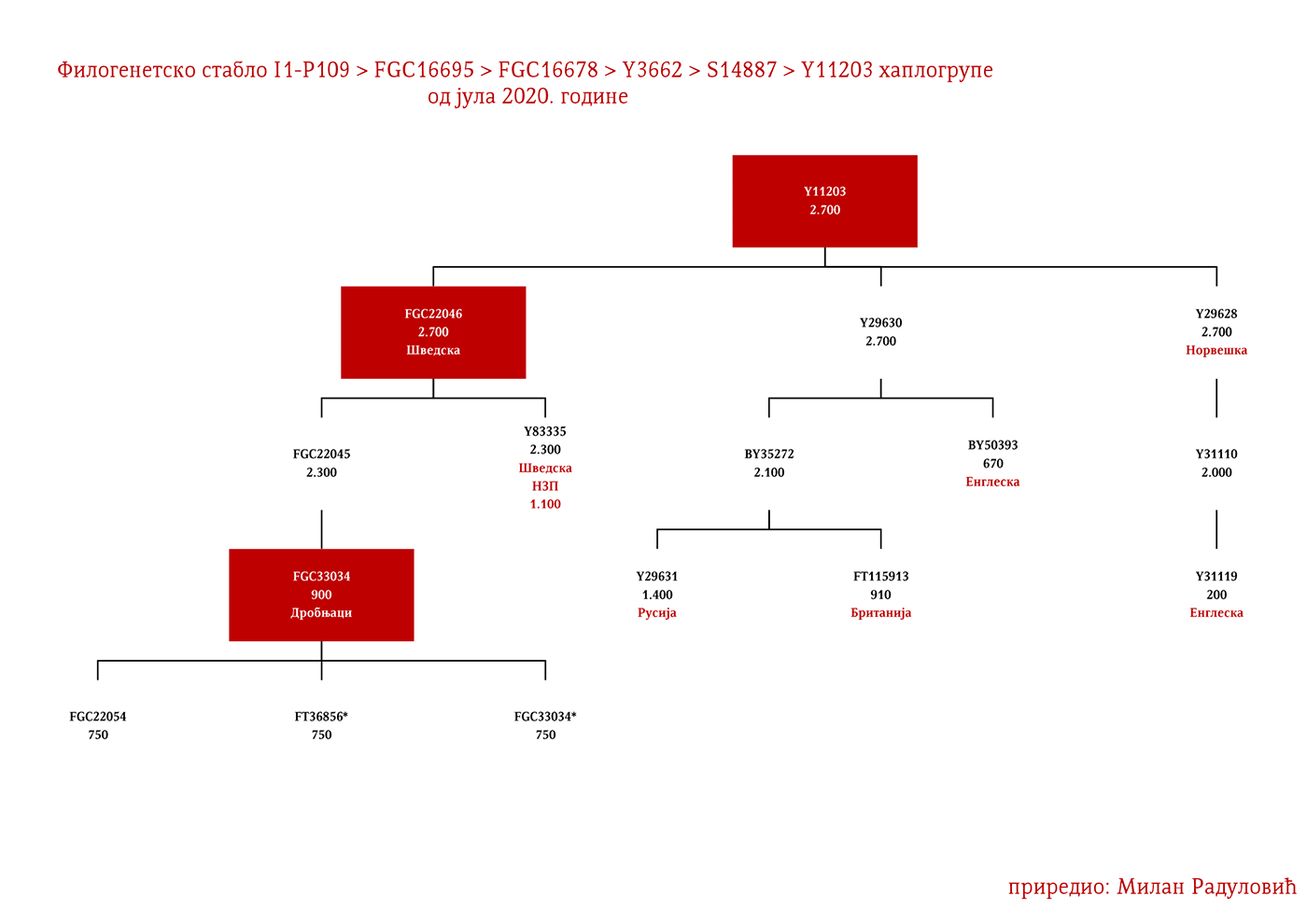
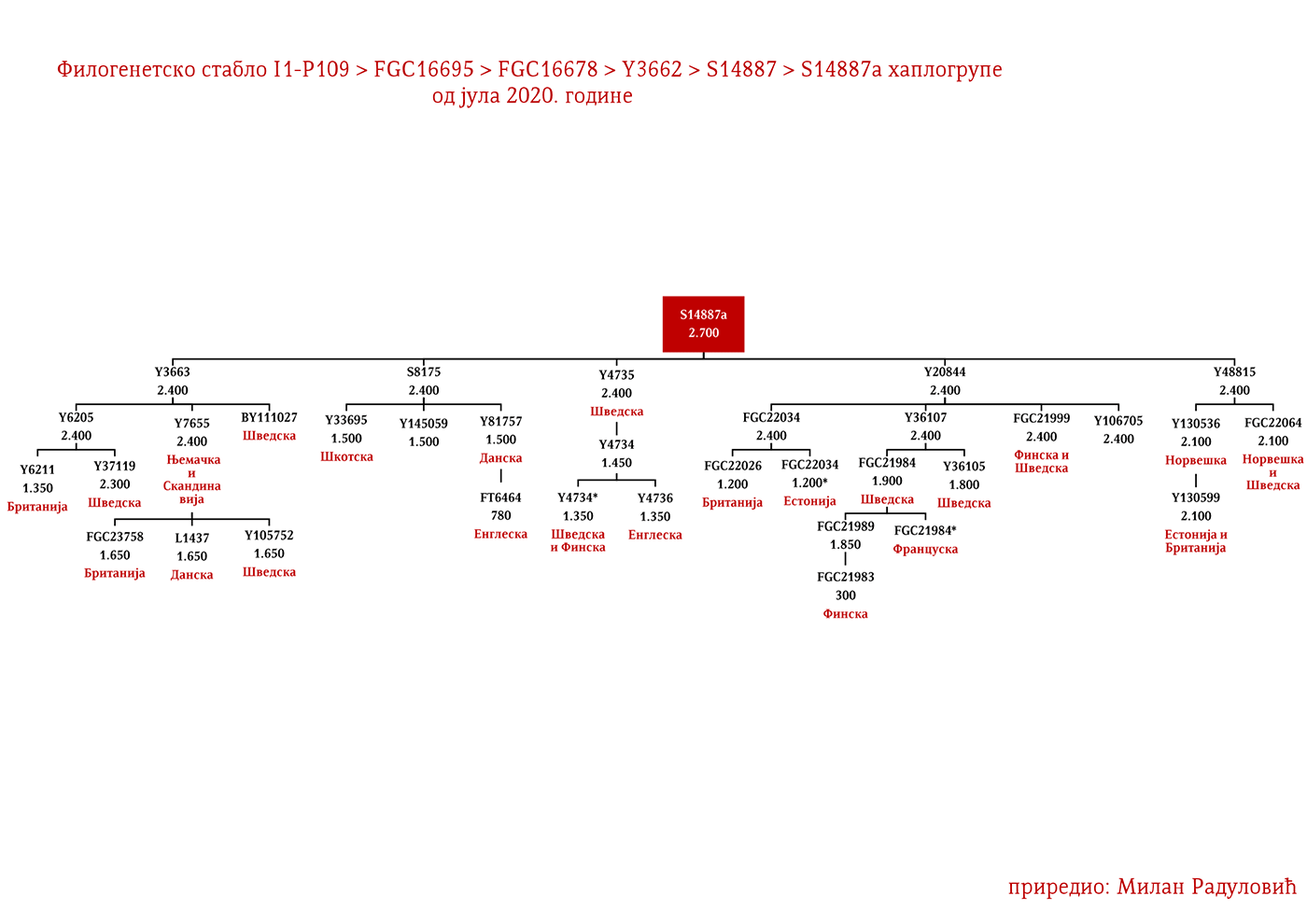
DROBNJACI I1-P109>FGC16695>FGC16678>Y3662>S14887>Y11203>FGC22046>FGC22045
Prvi pomen Drobnjaka je iz 1354. godine kada su braća Dabiživ, Bogoje i Hran, unuci Drobnjakovi, podigli crkvu u Vaganešu (Jireček, 1923, str. 54). O njihovom ranijem porijeklu ima više teorija baziranih što na predanjima što na konstrukcijama, stoga na ovaj dio starijeg porijekla odgovor daje genetička genealogija. Prema rezultatima do kojih smo došli ispitivanjem Y- DNK, a koji su javno poznati zahvaljujući Srpskom DNK projektu godinama unazad, Drobnjaci pripadaju I1-P109>FGC22045 haplogrupi. Od najbližih rođaka izvan Balkanskog poluostrva imaju Šveđane i druge nordijske narode. Po metodologiji starijeg grupisanja istraživača, njihov haplotip svrstava se u klaster poznat pod nazivom nordijski. Do vremena najbližeg zajedničkog pretka svih testiranih I1-FGC22045 muškaraca svega je 700-800 godina (YFull, 2020, pp. 8.06.01-8.09.00 ). Dakle, zajednički predak živio je negdje u 13. vijeku. Izvan prostora matične Stare Hercegovine i područja zemalja u kojima Srbi žive, njihove bliske srodnike srećemo u S. Makedoniji i na jugu Albanije te kod testiranog Rumuna (vjerovatno skorije asimilovana linija).
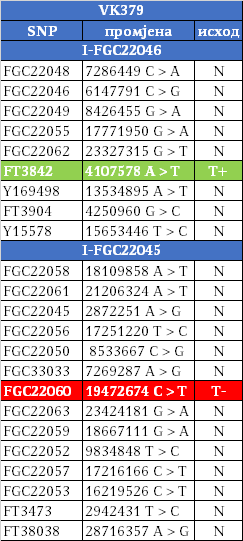
Kako su im najbliži Šveđani i drugi Nordijci, te kako izostaju u rezultatima drugih evropskih populacija i kako djele zajedničkog pretka u okviru 2200 godina sa nedavno otkrivenim Vikingom iz velikog arheogenetičkog istraživanja, VK379 iz Švedske, ovim potvrđujemo pisanje ranijih srpskih istraživača Siniše Jerkovića (Jerković, 2017), Jovice Krtinića (Krtinić, 2020) i drugih koji su pisali o vikinškom/normanskom porijeklu naših Drobnjaka. Dakle, svako povezivanje FGC22045 sa istočnogermanskim narodima za sada nema uporišta u rezultatima Y-DNK.
Kontakti Normana i Srba nisu istorijska nepoznanica. Skiličin Nastavljač početkom XII veka beleži podatak da je Bodinova vojska u Bici kod Prištine 1072. godine zarobila vizantijskog vojskovođu Longivardopula (grč. Λογγιβαρδόπουλος). Zarobljeni vojskovođa beše poslat knezu Mihailu u Zetu, koji se tako omili knezu, da mu ne samo slobodu vrati, već primivši od njega veru, da će ubuduće njemu služiti, dade mu svoju kćerku za suprugu. Longivardopul dobavi svojih zemljaka u pomoć, te kako beleži Skiličin Nastavljač, beše sastavljena nova vojska od srpskih i normanskih četa, koja pod vođstvom Longivardopula bi upućena u pomoć Bodinu u borbi protiv Vizantinaca (Rački, 1873).
Zanimljivo, kralj Mihailo će 1081. godine i svog sina Konstantina Bodina oženiti Normankom Jakvintom Barijskom. Podsećanja radi, iste godine su se Normani pod vođstvom Roberta Gviskara s 16000 ratnika iskrcali u Drač, gdje su se sukobili sa Romejima, a gdje su Srbi predvođeni Bodinom izbegli direktno učešće u bici gdje ih je pozvao romejski car (Živković, 2006). Da li je ova grupa ratnika ostala na prostoru srpskih zemalja nije poznato, kao što nije poznato ni šta se na kraju desilo s normansko-srpskom vojskom koju je predvodio Longivardopul, ali je evidentno da je jedan muškarac imao osnivački efekat nekih 100 godina kasnije od ovih dešavanja.
Po narodnom sjećanju koje su zapisali brojni istraživači, Drobnjaci su se po dolasku na Jezera sukobili sa starosjedilačkim plemenom Kriča. Borbe između njih bile su dugotrajne i iscrpljujuće. Kako su Kriči dali žilav otpor, Drobnjacima su u pomoć priskočila druga okolna plemena, te su Kriče potisli prema planini Kučajevici gdje su izazvali pomor među njima. Neko vrijeme nakon toga Kriči su se iskupili u Foči i preko Pive od strane Durmitora udarili na Drobnjake, ali su ih ovi dočekali. Borbe između Drobnjaka i Kriča bile su dugotrajne i iscrpljujuće, a krajnji rezultat borbi bio je prebacivanje Kriča na desnu obalu Tare (Luburić, 1930). Narodno predanje pripovjeda da je na drobnjačkom zemljištu počela borba protiv Grka, koja je kulminirala u doba Stefana Nemanje.
Tokom srednjeg vijeka bili su u vlaškom statusu, a prema njihovom predanju učestvovali su u sukobu Nikole Altomanovića i kneza Lazara Hrebeljanovića na kneževoj strani, predvodio ih je vojvoda Mileša Mandić nakon čega ih je knez Lazar darivao povlasticama; u Kosovskom boju predvodio ih je Đurjan Kosovčić (Luburić, 1930, str. 22, 95).
Iskopavanjem posmrtnih ostataka iz grobnice kneza Nikole Raškovića Drobnjaka, vlastelina iz 15. vijeka, u Cernici 2018. godine i obradom njegovog Y-DNK profila nepobitno smo utvrdili činjenicu da se radi o pripadniku drobnjačkog plemena. No, o knezu Nikoli biće više reči u knjizi „Genetičko poreklo Srba Stare Hercegovine”.
Tokom otomanske okupacije, migracioni pravci Drobnjaka išli su mahom u smjeru drugih dinarskih krajeva, tako ih srećemo zajedno sa Kričima i Pivljanima u Krajini. Snažno žarišno prisustvo roda u Polimlju ne čudi jer je ono naslonjeno na matičnu regiju, dok je jako prisustvo na Romaniji posledica migracija iz Drobnjaka tokom burnih godina 18. i 19. vijeka i nešto manje onih sa kraja 17. vijeka.
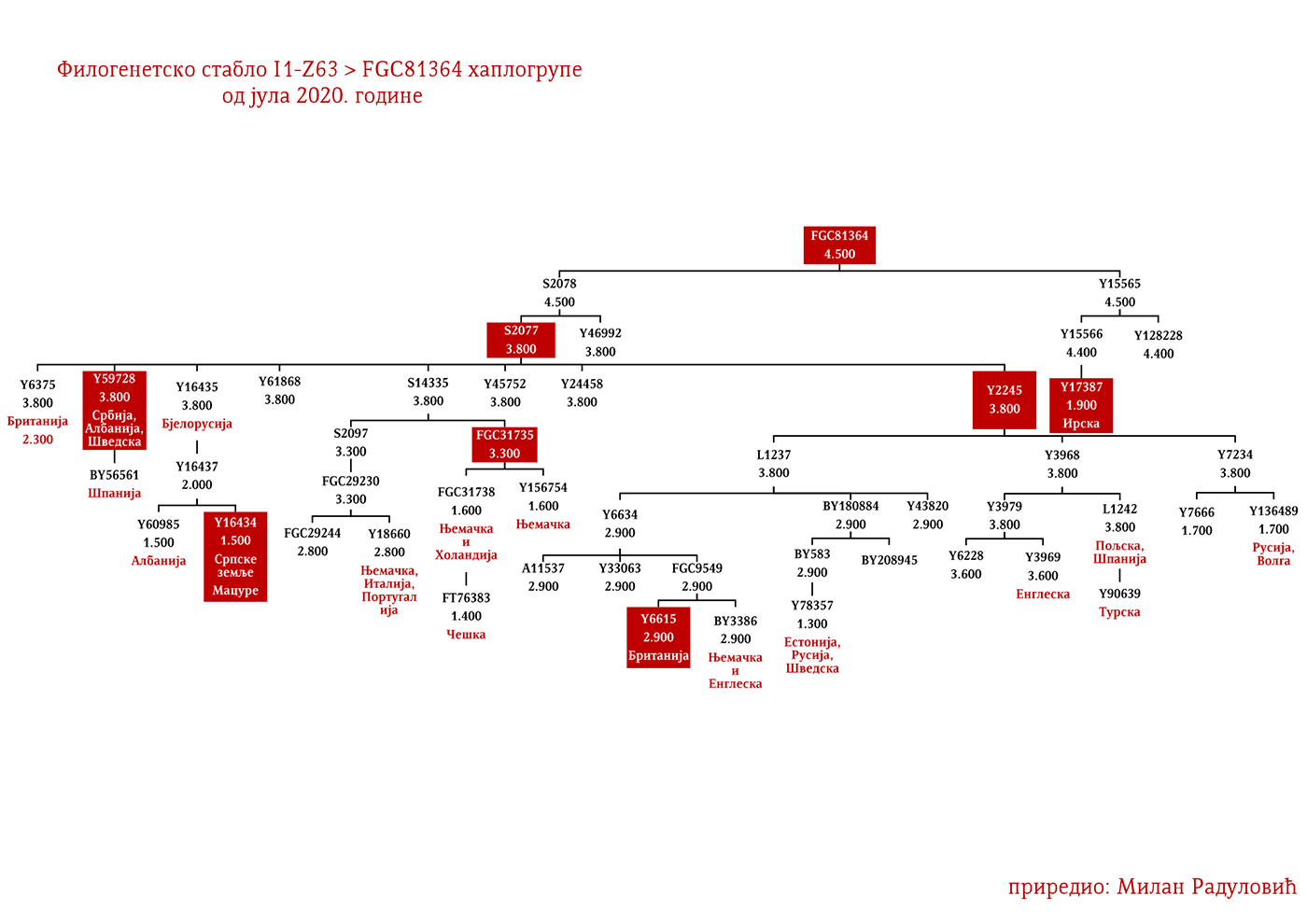
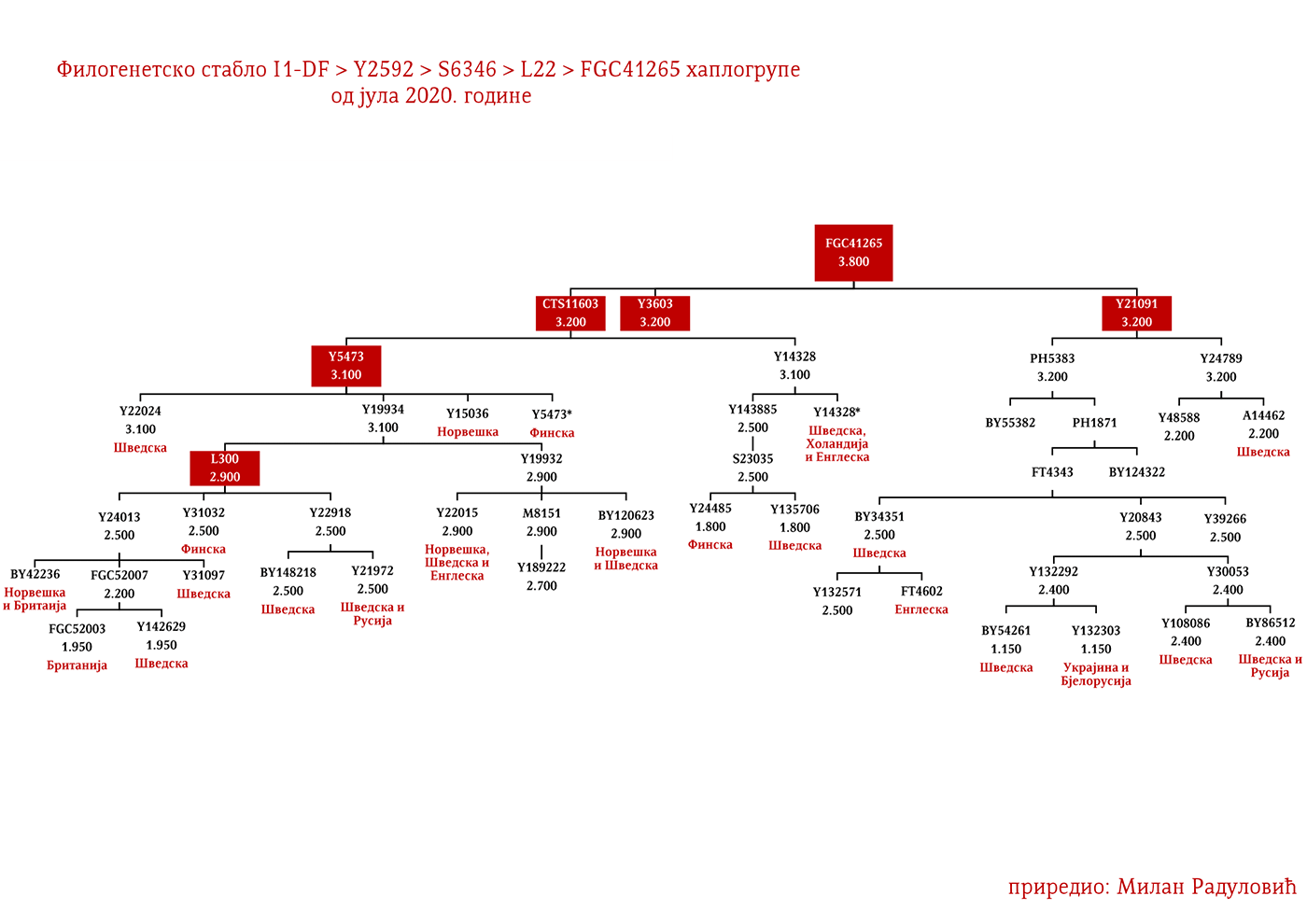
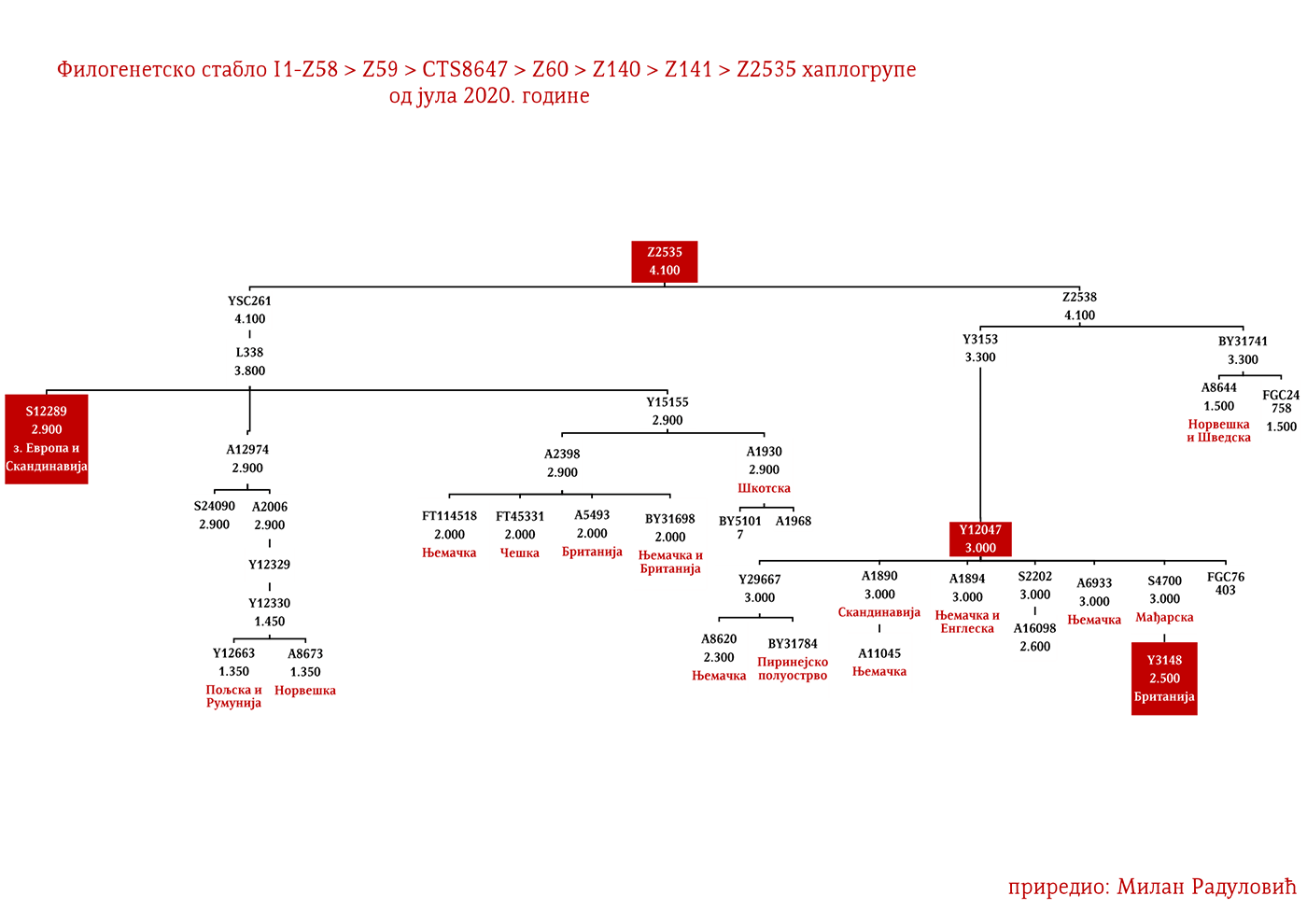
MACURE I1-Z63>FGC81364>S2078>S2077>Y16435>Y16437>Y16434
Macure su prema usmenoj tradiciji stari predslovenski narod sa prostora današnje Crne Gore, iz njenog istočnog dijela, a koji je živio na području današnjih Pipera, Bratonožića, Moračana, Rovčana i Vasojevića. Ostali su toponimi kao svjedočanstvo o ovom plemenu, i to su između ostalog, pašnjak Macurovine iznad sela Liješnje, dio šume Kunak u naselju Cerovica sa nazivom Macur brijeg; brojna stara groblja u selima Trmanju, Liješnju, Veljem Dubokom, Sreteškoj Gori, Međuriječju, Cerovici te u Gornjim Rovcima, gde se dva groblja nazivaju macurskim, ali i u brojnim drugim mestima (Vujanić, Jerković, Krtinić, & Marinković, 2014). Kako su svi toponimi Macura vezani za visoravni i planinske predjele, analogno zaključujemo da se radilo o stanovništvu koje se pretežno bavilo stočarstvom, odnosno koje je kasnije tokom srednjeg vijeka dolazilo iz vlaškog staleža.
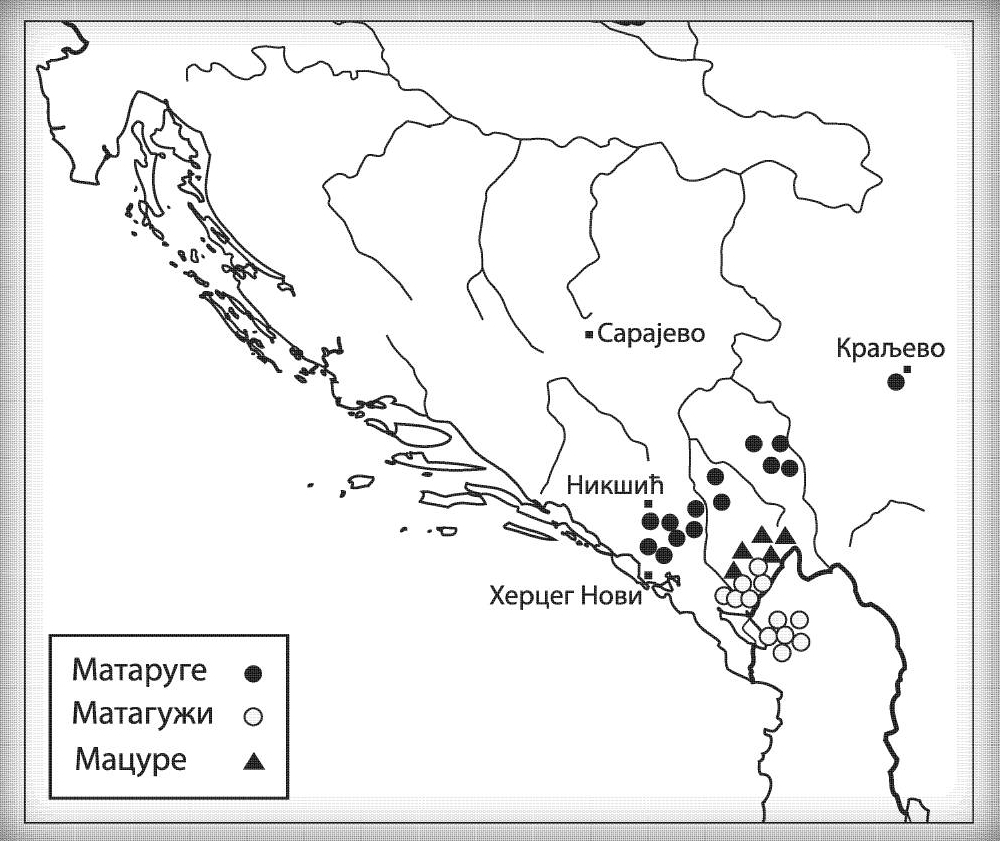
Kao razlog naglog nestanka ovog plemena navode se oštre zime, gladne godine i njihovi međusobni sukobi, te je tako u Crnoj Gori ostala uzrečica „poklali se k’o Macure“ ili „pobiše se k’o Macure“ (Vujanić, Jerković, Krtinić, & Marinković, 2014, str. 44). Putem Y-DNK analize nosilaca prezimena Macura iz Krajine i njima srodnih bratstava iz Brda u Crnoj Gori (v. Tabelu Srpskog DNK projekta), utvrdili smo praporijeklo ovog starog plemena, a ono je u najkraćem rečeno germansko i pripada haplogrupi I1- Y16434. Pregledom cjelokupne predačke Y16435 haplogrupe, najvjerovatnije se radi o liniji proistekloj od istočnogermanskih naroda.
DRUGI SRPSKI I1 RODOVI
Osim drobnjačke I1-P109 podgrane, na našim prostorima srećemo još najmanje dvije, a to su Y13930 i Y5621. Haplogrupu I1-Y13930 srećemo u dinarskoj zoni i njeno matično područje je istočna Hercegovina; dok je Y5621 prisutna u okolini Vranja i vrijedi pomenuti da ovoj grani pripadaju dva vikinška uzorka VK70 i VK281.
Rod koji za sada nije imenovan a koji pripada I1-Z63>Y13946>Y51867>PH220 haplogrupi rasut je širom srpskog etničkog prostora, od Rasine do Bosanske Krajine. Dalje rođake van našeg prostora srećemo u Bugarskoj na udaljenosti od oko 1200 godina, a paralelna bratska grana Y51867>S22866 prisutna je u Švajcarskoj i Njemačkoj, dok je zajednički predak Y51867 grane živio prije 2300 godina. Ravnomjernija rasprostranjenost nam govori u prilog ranom učestvovanju u etnogenezi Srba, dok u kombinaciji sa prethodno navedenim udaljenostima sa drugim Y51867 muškarcima izlačimo zaključak o predslovenskom porijeklu roda, odnosno vrlo vjerovatno se radi o potomcima germanskih plemena koja su prodrla na Balkan. Naravno moguće su i druge opcije doseljavanja u predslovenskom vremenu, no istakli smo ovu vjerovatniju.
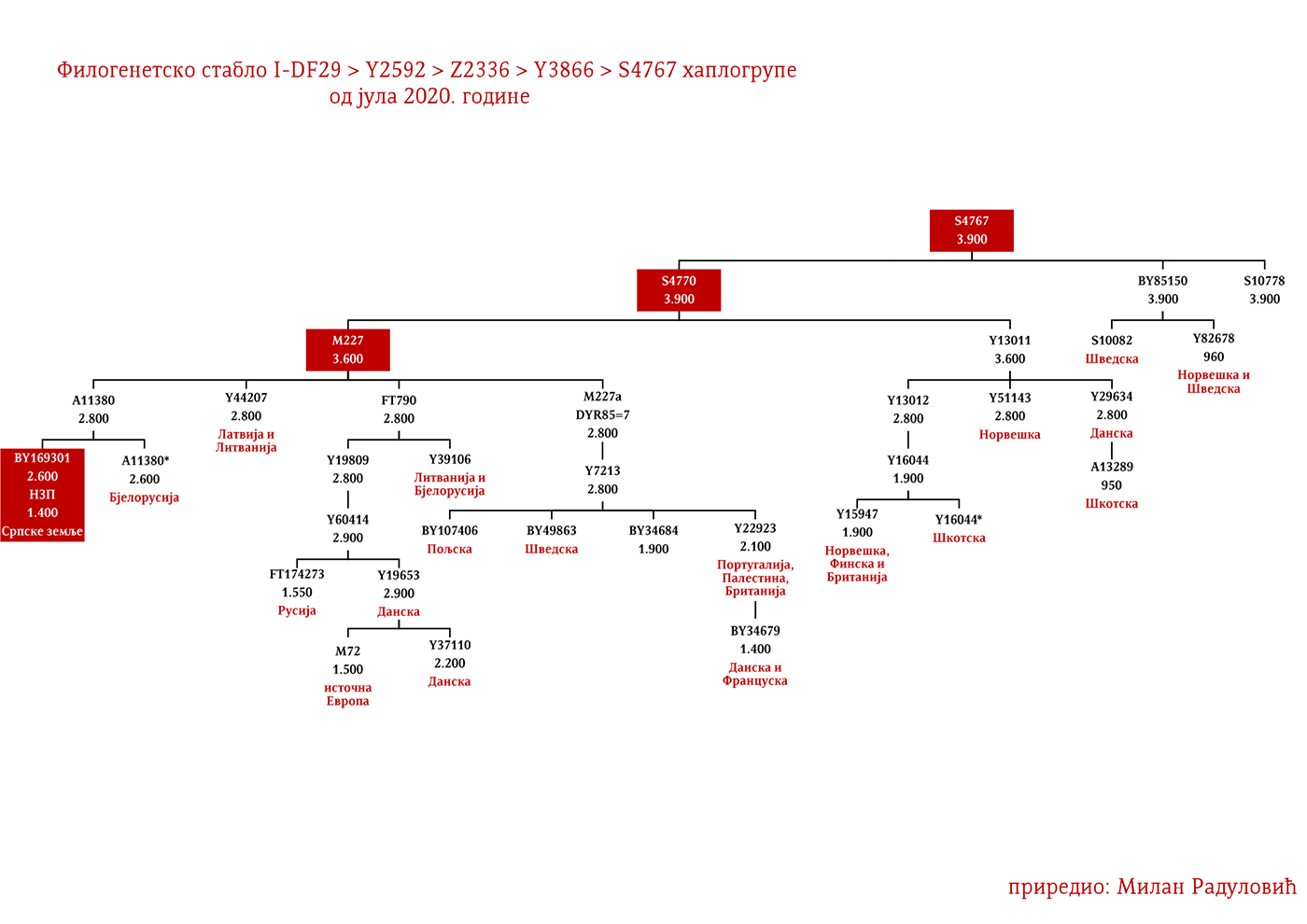
Još jedan rod koji je brojan a nije imenovan, pripada podgrani I1-M227>A11380>BY169301 i rasut je širom srpskog etničkog prostora, što nam govori o njihovom ranom učestvovanju u etnogenezi Srba. Izvan Balkana, u okviru od 2500 godina srećemo njihove srodnike u Bjelorusiji i Austriji. Moguće je da su gotskog porijekla.
Prisutne su i druge linije kod nas, ali kako nisu naročito zastupljene i kako nisu dovoljno profilisane o njima ne možemo nešto podrobnije pisati i izvlačiti zaključke.
POZNATE LIČNOSTI

BIRGER JARL
Bio je jarl Švedske (1248-1266) i zet švedskog kralja Erika XI. Dolazio je iz porodice Folkunga i osnovao je dinastiju Bjalbo. Predstavljao je jednog od najvećih švedskih državnika srednjeg vijeka. Testiranjem njegovih posmrtnih ostataka i njegovog sina ustanovljeno je da pripadaju haplogrupi I1 (Malmström, et al., Finding the founder of Stockholm, 2012).
NIKOLA RAŠKOVIĆ DROBNJAK
Srpski je vlastelin sa područja Gacka koji se pominje u dubrovačkim spisima 1453. godine. Tokom iskopavanja na lokalitetu pod Makovim valom u Cernici kraj Gacka 2018. godine, uz temelje crkve-zadužbine kneginje Jelene Lazarević nalazi se grobno mjesto kneza Nikole čiji su uzorci testirani u laboratoriji Biološkog fakulteta iz Beograda. Iskopavanje je vodio Muzej Hercegovine u Trebinju, dok je Društvo srpskih rodoslovaca „Poreklo“ bilo inicijator istraživačkog poduhvata. Prema rezultatima koje je obradio tim Srpskog DNK projekta, knez Nikola Rašković pripada haplogrupi I1-P109>FGC22045 i rodu Drobnjaka.

DINASTIJA GRIMALDI
Dinastija Grimaldi je italijanskog porijekla i vladala je u periodu od 1297. godine do 1731. godine, kada je umro posljednji član ove porodice. Nakon smrti posljednjeg člana dinastije Grimaldi na vlast dolazi dinastija Matinjon-Grimaldi koja je vladala do 1949. godine. Monakom danas vlada dinastija Polinjak-Grimaldi. Pripadaju haplogrupi I1-P109 (Grimaldi – Family Tree DNA, 2020).
PORODICA LI
Prema podacima sa projekta Li prezimenjaka (Lee Surname DNA Research Project, 2020, p. Subgroup 21: Lees of VA (Richard Lee m. Ann Constable), poznata porodica Li iz Virdžinije i Merilenda od kojih su bili Ričard Prvi Li, najveći veleposjednik u SAD-u i čuveni južnjački general Robert Li pripada I1-P109>S10891>Y13930>Y14227>Y14225>BY70495.

ALEKSANDER HAMILTON
Aleksander Hamilton (1757-1804) je jedan od „Očeva osnivača SAD“. Bio je američki političar, ekonomista i prvi sekretar finansija SAD. Zahvaljujući njegovoj zamisli je sazvana Filadelfijska ustavotvorna skupština, a bio je i koautor Federalističkih spisa, djela koje se smatra glavnim izvorom za tumačenje Ustava SAD. Prema rezultatima sa Hamilton DNK projekta (HAMILTON SURNAME DNA RESULTS AND DISCUSSION, 2020, pp. Group I1-5), Aleksandar je pripadnik I-Z59>CTS8647>Z60>Z140>Z141>Y15150>Y48678 haplogrupe.
ENDRU DŽEKSON
Džekson Endru Džekson (1767-1845) bio je američki predsjednik i osnivač današnje Demokratske stranke, takođe je bio i vojskovođa. Poznat je po tome što je bio 7. predsjednik SAD. Prema analizi genealogije Heja (Hay, Thread: President Andrew Jackson belonged to haplogroup I1, 2014) i rezultatima sa projekta Džeksona (The Jackson Project, 2014), on je pripadao I1 haplogrupi.

LAV NIKOLAJEVIČ TOLSTOJ
Lav Tolstoj (1828-1910) bio je ruski grof i pisac, jedan od velikana svjetske književnosti i jedan od najvećih realista toga doba. Poznat je po svoja dva najveća djela, „Ana Karenjina” i „Rat i mir”. Testiranjem njegovog potomka Petra Tolstoja utvrđeno je da pripada haplogrupi I1 (Hay, Origins and history of Haplogroup I1 (Y-DNA), 2016).
DRAGOMIR BOJANIĆ GIDRA
Dragomir Bojanić-Gidra (1933-1993) bio je srpski pozorišni, televizijski i filmski glumac. Glumio je u više od sto uloga u filmovima i TV serijama, a uloga Žike Pavlovića u filmskom serijalu „Lude godine” ostala je kao kultna. Šezdesetih godina prošlog vijeka u Italiji snimio je desetak vestern i akcionih filmova. Za ulogu u filmu Balada o revolverašu (1967) režisera A. Kaltabijana dobio je i nagradu. Pripada haplogrupi I1-P109>FGC22045 i rodu Drobnjaka.







-270x270.jpg)



















3. decembar 2020. u 12:44
Neko,tamo daleko...
Nisam genetičar ali jedna stvar mi nije jasna.Po kom kriterijumu je I1 starija od I2 haplogrupe? Tj. zašto je zapadna grana redni broj 1 a istočna redni broj 2? Procjena nastanka I1 je prije 27 500 godina.Informacija iz vašeg teksta.Okvirno 25 500 prije Hrista.Na zvaničnoj stranici Vikipedije procijenjena starost,tj nastanak I2 je između 26 000 i 31 000 stare ere.Matematika nam govori da je I2 stariji minimum 500 godina.Kao što sam napisao,nisam genetičar ali na prvu loptu nešto ne štima.Ili ste vi napravili grešku pa ste umjesto 27 500 prije Hrista napisali samo 27 500 predvidivši i 2020 nove ere ili ovdje postoji klasična diskriminacija Istoka Evrope od strane naučnih krugova na Zapadu,tj. isticanje svoje grane kao primarne a mrski istočnjaci po starom nepisanom pravilu sekundarna kategorija? Opet sve i da je I1 nastala oko 27500 godine stare ere ima li dokaza da je starija od I2 čija moguća starost ide i do 31 000 prije Hrista.Da se ne svodi na ono : njihovi instituti,njihova nauka,njihova pravila,njihova kategorizacija?
Gledajući tekst vidim i neke zanimljivosti.
Ekstremno mali broj testiranih u Norveškoj,Danskoj i Holandiji.Na prvi pogled mi deluje da što su ljudi bogatiji to ih manje zanimaju ovakve stvari…
Albanci su skočili sa 2.7 % 2010-te na nešto više od 4 % 2019-te (podatak sa njihovog DNK projekta). Da li je u pitanju neko štelovanje i manipulacija? To je značajan rast.
Iznenađujuće nizak procenat ove haplogrupe kod baltičkih naroda.Oni redovno ističu sebe kao “sinove Prusije”,tj u sekundarnoj varijanti nakon svog ugro-finskog porijekla.Tačno da se to prusko odnosi na društveni i kulturni aspekt ali realno je bilo zaočekivati značajniji procenat I1 na tom prostoru.Vrlo su blizu germanskoj kolevci.jugoizapadno Njemačka,severozapadno Skandinavija .Osim severnog kraja Estonije koji je u blizini Finske procenat ove haplogrupe zanemarljiv.
Rekao bih da je najveće iznenađenje upravo Srbija.Procenat od 8 % na tolikoj udaljenosti od matice je I1 zaista frapantan podatak.Ako može neko da mi odgovori iz administracije sajta ili neko ko bolje poznaje materiju kakva su dalja očekivanja tj . neke buduće projekcije zastupljenosti ove haplogrupe u Srbiji? .Smanjenje,povećanje ili ostanak na ovom procentu? Deluje mi na prvu loptu kao da su Drobnjaci i Hercegovci bili najzainteresovaniji za srpski DNK projekat…
3. decembar 2020. u 19:13
Branko Todorović
Ove brojke se odnose na pojedine radove koje je autor koristio. To svakako nije ukupan broj testiranih pojedinaca iz neke zemlje. Tako se moglo uzeti i neko istraživanje za Srbiju, BiH ili Crnu Goru, i napisati da je procenat uzet na osnovu uzorka od 180 ljudi. Takođe, kada je reč o udelu haplogrupe kod raznih naroda, treba imati na umu da nisu svi jednako brojni, kao i da raznovsrnost podgrana nije svuda jednaka. Kod nas je visok procenat, ali je mali broj pripadnika ove haplogrupe, kao što je mali i broj njenih podgrana, za razliku od skandinavskih zemalja ili Nemačke, gde je njihov broj višestruko veći, po oba parametra.
5. decembar 2020. u 00:26
Dubrovski
Uz svo duzno poštovanje za uloženo u ovu analizu, smatram da je prepostavka da su Drobnjaci Normani zaista smesna. Sami navodite kako je verovatno jedan od tih sto su dosli da pomognu Bodinu posle ostavio svoje tragove. I eto tako vec njegov unuk napravi brojno i mocno srpsko brdjansko pleme. Koi je će to Švedjo ??najamnik da se skrasi u nasim gudurama??? Dajte molim vas.
Normani su uglavnom bili Danci. Veća je verovatnoća da je to gotska grupa, mada i to teško , jer te grupe nema u Španiji , Italiji, Krimu, gde su oni otišli i ostali, sigurno u vecem broju nego na Balkan i sa vise motiva da odu tamo no da ostanu odje.
Baltički narodi su u velikoj vecini R1a, tako i , od Nemaca i Poljaka istrebljeni Prusi. Jako je zanimljivo i za veliku sumnju da su istočnogermanska plemena tako ekspanzivna i vojno napredna naprasno nestala i prepustila celu Istočnu Evropu , pa i pola Njemačke miroljubivim Slovenima iz močvara Pripjata, koji su, gle čuda za sto godina okupirali i do dan danas zadržali svoje ime u čitavoj Istočnoj Evropi.
Srdačan pozdrav
5. decembar 2020. u 00:30
Dubrovski
Koja je haplogrupa Varjaga? R1b ili I1? slabo su nešto ostavili traga ti Švedjani po Ukrajini i Rusiji?
Varjag je slovenski izgovor švedskog narodnog imena SVERIGE
9. decembar 2020. u 12:13
Goran
Narod koji je takođe dugo vremena boravio na Balkanu i koji veoma mnogo uticao na istorijska dešavanja na jugu Evrope jesu Goti.Goti su svakako ostavili i značajan genetski trag na Balkanu sobzirom dasu i Vizigoti dugo vremena boravili u Dakiji i Trakiji,išli do Epira a sa krajnjeg juga Vizigoti su otišli preko Dalmacije do Sev.Italije i dalje na Zapad.Ostrogoti su boravili u Panoniji odakle su otišli u Prevalis(Duklja,Zeta)odakle odlaze u Italiju a zatim formiraju svoju kraljevinu koja obuhvata Italiju i svu Balkansku Dalmaciju.Njihova kraljevina na Balkanu je trajala oko 50godina ai posle poraza Ostrogota od Vizantinaca,mnogi Ostrogoti na Balkanu nisu trenutno nestali već su mnogi ostali u brdima Balkana u 5 i 6veku gde su dočekali Slovene.Neki su bili poluromanizovani stočari planinci nazvani Vlasi,a neki su u 6,7veku bili Slovenizovani.U genetskom smislu su za Srbe i druge Balkanske narode veoma bitni jer su dugo vremena bili prisutni na Balkanu skoro tri veka,bili su poreklom iz Skandinavije iz Gotlanda i otišli su daleko na istok prema Crnom Moru tipični I1Z63.Još dok su bili u zajednici kao Goti pre podele na Ostrogote i Vizigote dok su živeli na ušću Visle tu su boravili zajedno sa Burgundima koji su takođe poreklom Skandinavci Angli sa ostrva Borholm.Goti su poveli jedan broj Burgunda sa sobom prema reci Odri gde susreću Vandale.Sa Gotima idu i Gepidi i u blizini Crnog mora susreću Germanske Bastarne i Skire.Takođe na reci Odri mogli su im se pridružiti i brojni Rugijci.Tako su Goti kao narod došli na Balkan sa većim brojem Germanskih haplogrupa podgrana vodeći sa sobom pripadnike većeg broja Istočnih Germanskih plemena koja svako ima svoju genetsku podgranu.Tako napr.Goti su asimilirali jedan broj Burgunda još na ušću Visle,koji su već u 4veku bili potpuno integrisani u Gote i Istočne i Zapadne.Burgundi bi mogli da budu nosioci I1Z63-51867 i ovi Goti-Burgundi su mogli tu podgranu da donesu iu Bugarsku i Makedoniju,Srbiju,Hercegovinu,Dalmaciju,tako iu Italiju,Španiju,gde su sve išli Goti.Takođe Goti su sobom vodili i grupe Vandala,Gepida Rugijaca koji su Gotima doneli I1Z63-16435 koje su sa Gotima doneli na Balkan.Goti su sobom vodili i grupe Bastarna i Skire koji su Gotima preneli I2M227 koju su kao Goti doneli u Dalmaciju.Sami Goti kao Skandinavci su mogli da budu nosioci I1Z63-L22 koja je tipična za Skandinavce a koja je dosta brojna u Staroj Hercegovini pored drugih Gotskih podgrana (I1Z63-51867-Ph220),I2M227, I1Z63-16435,što su sve imala Germanska plemena koja su asimilirali Goti na svom putu od Skandinavije ka južnoj Evropi.Ove Gotske,,Germanske,,haplogrupu-podgranu su dokaz dasu mnogobrojni Goti ostali i bili Slovenizovani na Balkanu.
7. mart 2021. u 17:58
Goran
Mnogi rodovi koji imaju aplogrupa I1Z63 svoje poreklo vezuju za Staru Hercegovinu,Nikšić,Staru Župu Onogošt i Grahovo.Mnogi rodovi koji su I1Z63 su iz Onogošta u srednjem veku otišli prema Dubrovniku,Trebinju,Kotoru,Popovom polju,Gacku.Skoro svi I1Z63 u Staroj Hercegovini mogu se povezati sa Starim Onogoštom,Nikšićkom poljem i tvrđavom Anagastum.Ovu tvrđavu u 5.veku su podigli i u njoj iu polju oko nje,živeli Ostrogoti.U vreme doseljavanja Slovena u širu okolinu tvrđave Anagastum u 7veku,potomci Ostrogota iz Anagastuma ostali su u poljima i župi pored tvrđave,gde su se sjedinili sa doseljenim Slovenima R1a.Sobzirom na visok procenat I1Z63 haplogupe kod rodova koji su poreklom od Onogošta-Nikšića-Riđana,može se zaključiti daje veći roj Ostrogota ostao u brdima i poljima oko Anagastuma gde su dočekali Slovene koji su tvrđavu nazvali Onogošt,i tu formirali svoju Župu.Verovatno da između Ostrogota i Slovena u Onogoštu nije bilo nikakvih sukoba,već su Ostrogoti bili dobrovoljno vrlo brzo Slovenizovani i ostali da kao Sloveni žive u Župi.Možemo slobodno reći da su Srbi sa haplogupom I1Z63, poreklom od Nikšića,potiču od Slovenizovanih Ostrogota iz Anagastuma,tvrđave sa granice Prevalisa.