



Српски ДНК пројекат је покренуо објављивање серије прилога у којима читаоце упознаје са основама генетичке генеалогије, дисциплине која последњих година наилази на све веће интересовање. Серијал је настао као допуњена и прилагођена верзија чланка „Генетика у служби родословља“ (В.Михић, М. Рајевац, С. Јерковић), објављеног у књизи „Генетичко порекло Срба Старе Херцеговине“ (2021). Оваква врста садржаја до сада није била доступна на српском језику у форми научно-популарне грађе онима који су желели да се упознају са основама ове нове научне области. Зато смо припремили „Малу школу генетичке генеалогије“ кроз коју ћемо дати јасна објашњења и примере који приближавају појмове и методе савремене генетике, као и њихову примену у истраживању порекла породица, родова и народа.
У првом наставку серијала представили смо основне појмове генетике: шта су и како функционишу гени и ДНК, шта су хромозоми и како се одвија репродукција и наслеђивање. Било је речи и о људском геному, генетичким мутацијама, као и о STR и SNP маркерима који чине основу савремених генетичких тестова.
Други наставак садржи објашњење врста и намене генетичких тестова, као и појам хаплогрупа. Читаоци су упознати са филогенетичким стаблима и методама одређивања припадности Y‑DNA хаплогрупи, уз пример како се резултати тумаче и повезују са родословним истраживањима.
У трећем наставку који следи дајемо преглед основних Y‑DNA хаплогрупа у свету. Биће речи о њиховој старости, географској распрострањености и значају у историјском контексту, као и о хаплогрупама заступљеним код Срба.
ПРЕГЛЕД ОСНОВНИХ Y-DNA ХАПЛОГРУПА
Овде ћемо дати кратак преглед свих основних Y-DNA хаплогрупа у свету, заснован на YFull стаблу и ISOGG класификацији. Слика која следи приказује гранање Y-DNA хаплогрупа на филогенетичком стаблу мушке популације света. На графикону се може видети време издвајања сваке од основних хаплогрупа, а почетак ширења усправних линија у “троуглове” представља време када се процењује да је живео најскорији заједнички предак свих до сада тестираних припадника сваке појединачне хаплогрупе.
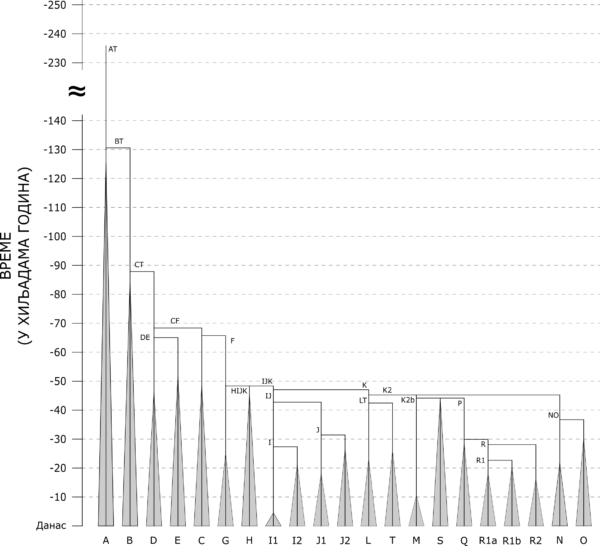
Хронологија гранања и експанзије основних грана филогенетичког стабла Y-DNA хаплогрупа[1]
Хаплогрупа А је најстарија хаплогрупа, у смислу да је то предачка Y-хаплогрупа целокупне данашње људске популације, са процењеним временом настанка пре око 235.000 година.[2] Иако се најчешће води као јединствена хаплогрупа, у стварности се састоји од чак четири хаплогрупе, редом А00, А0, А1а и А1b1, то јест све оне које не потпадају под макрохаплогрупу BT, од које потиче preko 90% данашњих мушких линија и која је настала од хаплогрупе А1b паралелно са сестринском хаплогрупом А1b1. Просторно су све четири А хаплогрупе скоро у потпуности ограничене на Африку, с тим што се поједине гране А1а и А1b1 срећу и на подручју Блиског истока и Европе. Највећи проценти забележени су међу којсанским народима Јужне Африке, Бушманима и Хотентотима,[3] као и међу говорницима нило-сахарских језика, попут народа Динка и Шилуки из Јужног Судана[4].
Хаплогрупа В издвојила се из макрохаплогрупе ВТ пре око 88.000 година.[5] Као и хаплогрупа А, скоро у потпуности је ограничена на афрички континент, с тим што се у малом проценту среће и на подручју Блиског истока и централне Азије (Арабијско полуострво, Иран и Авганистан). Највеће проценте бележи у појединим пределима централне и западне Африке, пре свега међу пигмејским народима.[6]
Хаплогрупа D настала је пре око 65.000 година у Африци, издвајањем из хаплогрупе DE.[7] Једна је од хаплогрупа које су биле део првих људских популација које су напустиле афрички континент и започеле насељавање Азије и Европе. Дели се на хаплогрупе D1 и D2. Грана D2 је веома ретка, засад је пронађена само код неколико појединаца из Африке (Нигерија) и са Блиског истока (Сирија и Саудијска Арабија).[8] Најбројнија и најраширенија грана је D1, чије су подгране прилично јасно раздељене у неколико региона. Најзаступљенија је код староседелачких народа Аину и Рјукјуанаца,[9][10] на Тибету[11], те код абориџинских племена Андаманских острва у Индијском океану.[12]
Хаплогрупа Е настала је пре око 65.000 година у Африци[13], где и дан данас бележи највише проценте. Има велики број старих и широко распрострањених грана од којих су најзаступљеније E-M2 у западној, централној и јужној Африци међу говорницима нигер-конгоанских језика (Банту), E-M81 на сeверу Африке код берберских народа, E-M123 на Блиском истоку и E-M78 на истоку Африке код кушитских народа. Огранак хаплогрупе E-M78 је и изразито европска хаплогрупа E-V13, која је најприсутнија у југоисточној Европи.
Хаплогрупа C настала је пре око 66.000 година из хаплогрупе CF.[14] Уз хаплогрупe D и F била је део најранијих људских популација које су напустиле афрички континент, и пронађена је у неким од до данас најстаријих тестираних људских скелета.[15] Иако се не може похвалити великом бројношћу, једна је од најраширенијих хаплогрупа у светским оквирима, чије гране се срећу од аустралијских Абориџина[16] и Папуанаца[17], преко Индијског потконтинента, Кине, Јапана, Сибира и централне Азије, па све до Блиског истока и Европе. Хаплогрупа C је карактеристична хаплогрупа монголских народа[18].
Макрохаплогрупа F је, као и хаплогрупа C, проистекла из CF пре око 66.000 година.[19] Макро у називу носи због чињенице да је из ње настао велики број данас најбројнијих и најраширенијих хаплогрупа, и ако је посматрамо као целину, она је убедљиво најбројнија хаплогрупа у светским оквирима.
Хаплогрупа G настала је пре око 48.000 година[20] и проистекла је из макрохаплогрупе F, највероватније негде на подручју Блиског истока. Карактеристична је за популације југозападне Азије, пре свега Кавказа и Анадолије, мада се јавља и у подручју Леванта, Ирака и Ирана, и даље на исток све до централне Азије.[21] У Европи се спорадично јавља у ниским процентима од 2-5%, а највеће присуство бележи у региону Алпа (10-15%) и на Сардинији (15-30%).[22] Може се повезати с доласком земљорадње с подручја Месопотамије и Анадолије у Европу и на Кавказ, јер је била убедљиво најбројнија хаплогрупа међу до сада тестираним скелетима неолитских земљорадника.[23]
Хаплогрупа H настала је пре око 48.500 година[24], највероватније негде на простору југозападне Азије или Ирана. Највећи проценат данашњих носилаца ове хаплогрупе припада грани H1, која је сконцентрисана пре свега на Индијском потконтиненту, где бележи и највеће проценте међу дравидским популацијама јужне Индије. У Европи хаплогрупа H1 je доминантна код ромске популације[25]. Грана H2 је данас изузетно ретка, међутим у прошлости је била прилично заступљена код неолитских популација Леванта, Анадолије и Европе.[26]
Хаплогрупа I настала је пре око 43.000 година из хаплогрупе IJ.[27] Данас је једна од најмногобројнијих и најраширенијих хаплогрупа на европском континенту и вероватно најстарија хаплогрупа настала у Европи. Пронађена је у скелетима носилаца граветијенске културе, једне од најранијих скупина људских популација које су насељавале Европу у периоду палеолита.[28] Дели се на две основне хаплогрупе, I1 и I2. I2 данас постиже највећу концентрацију у југоисточној Европи, посебно код Срба, Хрвата и Бошњака, док је I1 доминантна у северној Европи (Скандинавија).
Хаплогрупа J настала је пре око 43.000 година из хаплогрупе IJ[29], највероватније негде на простору који обухвата источну Анадолију, јужни Кавказ, западни Иран и северну Месопотамију. Дели се на две основне хаплогрупе, Ј1 и Ј2. Хаплогрупа Ј1 високе проценте бележи међу семитским народима, Арапима и Јеврејима, пре свега на Арабијском полуострву и Леванту, али и широм Блиског истока и северне и североисточне Африке.[30] Друго подручје високе концентрације ове хаплогрупе је Кавказ, где су највиши проценти забележени код североисточних кавкаских народа.[31] У Европи су највиши проценти ове хаплогрупе забележени код острвских популација Крита и Малте.[32] Хаплогрупа J2 карактеристична је за данашње популације Анадолије, Кавказа и Медитерана. Њено присуство у Европи често се повезује са ширењем земљорадње током неолита. Дели се на две основне гране, Ј2а и Ј2b.
Макрохаплогрупа K настала је пре око 47.200 година[33], а из ње је простекао велики број неких од данас најбројнијих и најраширенијих хаплогрупа. Из њене гране К1 проистекле су хаплогрупе L и Т, из гране К-M2335 хаплогрупе N и O, а из гране К2b хаплогрупе M, S, Q и R.[34]
Хаплогрупа L настала је пре око 42.600 година[35] издвајањем из хаплогрупе LT, највероватније негде на простору југозападне или централне Азије. Дели се на две основне гране, L1 и L2. Грана L2 је веома ретка и пронађена је код свега неколицине појединаца у свету. Грана L1 даље се дели на L1b и L1a.[36] Највећа концентрација хаплогрупе L забележена је у јужној и централној Азији, пре свега у Шри Ланки, Пакистану, Авганистану, Индији и Ирану. У наведеним регионима ради се скоро у потпуности о грани L1a.[37] У Европи, на Кавказу и на Блиском истоку најчешћа грана хаплогрупе L је L1b.[38]
Хаплогрупа Т је сестринска хаплогрупи L јер је такође проистекла из хаплогрупе LT пре око 42.600 година.[39] Иако је раширена на веома великом простору, од западне Европе до Рога Африке и Индије, прилично је ретка. Високе проценте бележи једино код појединих афричких народа.[40] Пронађена је у скелетима неолитских земљорадника с подручја Леванта и Европе.[41] У Европи је присутна у ниским процентима, али се јавља у археогенетским налазима неолитских земљорадника.
Хаплогрупа М настала је пре око 44.300 година издвајањем из хаплогрупе К2b1.[42] Ограничена је скоро у потпуности на Папуу Нову Гвинеју, где бележи и највише проценте међу домородачким популацијама, док су мањи проценти забележени и међу популацијама околних острва Меланезије и Индонезије[43] и међу аустралијским Абориџинима.[44] Пример је „локалне“ хаплогрупе, ограничене на један географски регион.
Хаплогрупа S настала је као и хаплогрупа М, из хаплогрупе К2b1 пре око 44.300 година.[45] Такође је највећим делом ограничена на домородачке популације Папуе Нове Гвинеје[46], али се јавља у значајним процентима и код појединих популација Океаније и југоисточне Азије.[47]
Макрохаплогрупа P проистекла је из макрохаплогрупе K2b пре око 44.300 година[48], а из ње су настале хаплогрупе Q и R. Пронађена је у палеолитским скелетима из северног Сибира старим преко 30.000 година, који су, могуће, припадали предачкој популацији од које потиче значајан део данашњих популација Сибира, централне Азије и Европе.[49]
Хаплогрупа Q настала је издвајањем из хаплогрупе P пре око 30.000 година[50], највероватније негде у јужном или централном Сибиру. Заступљена је претежно код сибирских и централноазијских популација[51] и убедљиво је најзаступљенија хаплогрупа америчких Индијанаца.[52]
Хаплогрупа R настала је из макрохаплогрупе P пре око 30.000 година, највероватније на простору Сибира или централне Азије.[53] Хаплогрупа R дели се на млађе хаплогрупе R1 и R2, а R1 се дели на још млађе R1a и R1b. Гране R1a и R1b су данас најзаступљеније у Европи и чине основу већине европских мушких линија. Хаплогрупа R1a карактеристична је за евроазијски простор, од Европе преко Блиског истока и средње Азије, до Индијског потконтинента и Сибира. Грана R1а-М417, којој припада огромна већина свих данашњих носилаца R1а, по тренутно доступним резултатима одиграла је једну од кључних улога у настанку и ширењу индоевропских народа.[54] Хаплогрупа R1b настала је када и сестринска хаплогрупа R1a, пре око 23.000 година[55], такође негде на простору јужног Сибира или централне Азије. Врло рано је започела ширење на запад, јер се у Европи среће већ код једног скелета из периода палеолита.[56] Дели се на велики број млађих грана, од којих је данас убедљиво најбројнија и најраширенија грана R1b-M269, која се уз горепоменуту грану R1a-M417 сматра кључном у настанку и ширењу најранијих индоевропских племена. Од осталих грана треба поменути R1b-V2219, грану која је пронађена у мезолитским скелетима културе Лепенског вира[57], а која се данас у Европи може пронаћи само у траговима, док, изненађујуће, највише проценте бележи код појединих племена с подручја Камеруна и Нигерије (преко 90%)[58], који су највероватније последица неолитске сеобе из Европе преко северне Африке.[59] Хаплогрупа R2 настала је пре око 28.000 година издвајањем из хаплогрупе R.[60] Претежно је сконцентрисана на јужну и централну Азију, а највећи проценат забележен је код народа Бурушо у северном Пакистану.[61]
Хаплогрупа N настала је из хаплогрупе NO пре око 37.000 година[62], највероватније негде на простору (југо)источне Азије, где показује највећу разноврсност хаплотипова.[63] Највише проценте бележи на простору северне Евроазије, док се у мањем проценту јавља и у источној и централној Азији, и на Балкану.[64] Дели се на две основне млађе гране, N1 и N2. Грана N1 је убедљиво бројнија и раширенија грана, која се среће од југоисточне Азије, преко Сибира, све до северне и североисточне Европе. Кључна је за уралске народе и често се повезује са ширењем уралских језика. На простору Евроазије највеће проценте бележи код припадника уралске (Финаца, Хантија, Мансија, Ненеца и Нганасана) и турске језичке фамилије (Јакути, Долгани, Какаси, Туванци, Татари и Чуваши), као и код Бурјата, Евена, Евенка и Јукагира.[65] Грана N2 је веома ретка у светским оквирима, присутна је у малом проценту у Европи, а највећи проценат достиже управо код Срба (око 3%).[66]
Хаплогрупа O настала је такође из хаплогрупе NO пре око 37.000 година, на подручју источне или југоисточне Азије, где је и данас најзаступљенија.[67] У мањем проценту је заступљена и у Океанији, јужној Азији и на Мадагаскару. Дели се на две млађе гране, O1 и O2. Грана О1 највише проценте бележи међу становницима Јапана[68] и Јужне Кореје[69], и међу народима аустроазијске[70] (југоисточна Азија, источна Индија и Бангладеш) и аустронезијске језичке фамилије[71] (Тајван, Мадагаскар, Индонезија, Филипини и Океанија). Грана O2 је најзаступљенија код Хан Кинеза (преко 50%) и њима сродних народа из тибетанско-бурманске језичке фамилије с подручја Тибета, Непала и Бурме, али и код многих околних народа, Манџураца, Монгола, Корејаца, Вијетнамаца и других.[72] То је најбројнија хаплогрупа на свету по броју носилаца.
* * * * *
Све основне хаплогрупе се даље се гранају на огроман број млађих грана и подграна формирајући Y-DNA филогенетичко стабло човечанства. Откривањем нових СНП варијација из којих настају нове хаплогрупе, ово стабло се сваког дана допуњује и проширује. У новембру 2025. године на Y-DNA филогенетичком стаблу компаније YFull било је 64.704 грана, односно хаплогрупа, које су биле дефинисане са 429.611 СНП варијација.[73] У исто време Y-DNA стабло које одржава компаније Family Tree DNA садржало је преко 98.000 грана (хаплогрупа) и више од 837.000 СНП варијација.[74]
ЗАКЉУЧЦИ
Мушке популације већине светских народа, поготово европских, састоје се од великог броја различитих хаплогрупа, које су заступљене у већем или мањем проценту. Хаплогрупе се не могу повезати с нацијама, јер је припадност одређеној хаплогрупи биолошка (непроменљива) категорија, док су народи и нације друштвене (и самим тим променљиве) категорије.
Примена метода генетичке генеалогије за истраживање порекла породица и народа могућа је само уколико постоји размена генетичких резултата и ако се они међусобно упоређују и повезују. Зато су се већ од почетка комерцијалних тестирања почеле формирати базе генетичких резултата, родословни или национални пројекти. При томе је квалитет сазнања које такве базе дају у директној вези с величином базе. Сваки тестирани појединац својим резултатом доприноси употпуњавању и сагледавању целокупне генетичке слике појединих региона, народа, родова. Како би се у светлу генетичких резултата боље сагледала етничка прошлост српског народа, формиран је национални Српски ДНК пројекат, с циљем да на једном месту сакупи генетичке резултате Срба, али и народа с којима су Срби у прошлости делили простор и историју. Почевши са скромних око две стотине тестираних у 2012. години, када је основан, Српски ДНК пројекат је до данас израстао у један од највећих националних генетичких пројеката у Европи, који у својој бази тренутно садржи генетичке резултате преко 11.000 тестираних појединаца.[75] Постојање такве базе омогућава да се резултати ДНК тестирања сагледају и обраде на исправан начин и сместе не само у национални, српски, већ и у европски и светски генетички контекст. Могућности које пружа база Српског ДНК пројекта помажу да се тестирани појединци из регионалних истраживања групишу у родове по свом патрилинеарном пореклу, као и да се ти родови сигурније профилишу.
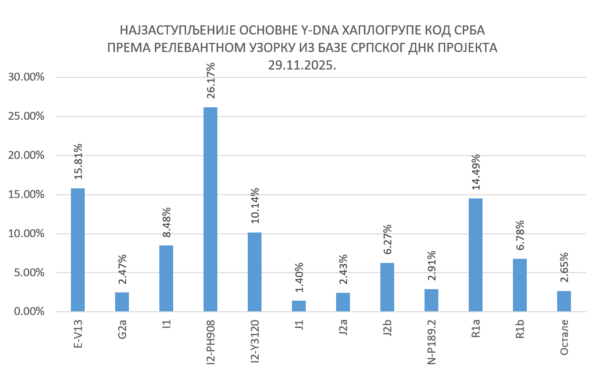
За крај, наводимо пресек заступљености Y-DNA хаплогрупа код Срба на основу података из базе Српског ДНК пројекта:
[1] Подаци о старости грана и времену живљења најмлађег заједничког претка (TMRCA) преузети са https://www.yfull.com/tree/A0-T/ (приступљено 29. 11. 2025).
[2] https://www.yfull.com/tree/A0-T/
[3] King TE, Parkin EJ, Swinfield G, et al. (March 2007). “Africans in Yorkshire? The deepest-rooting clade of the Y phylogeny within an English genealogy”. Eur. J. Hum. Genet. 15 (3): 288–93.
[4] Hassan HY, Underhill PA, Cavalli-Sforza LL, Ibrahim ME. Y-chromosome variation among Sudanese: restricted gene flow, concordance with language, geography, and history. Am J Phys Anthropol. 2008 Nov;137(3):316-23. doi: 10.1002/ajpa.20876. PMID: 18618658.
[5] https://www.yfull.com/tree/B/
[6] Underhill PA, Shen P, Lin AA, et al. (November 2000). “Y chromosome sequence variation and the history of human populations”. Nat. Genet. 26 (3): 358–61.
[7] https://www.yfull.com/tree/D/
[8] Haber; et al. (2019). “A Rare Deep-Rooting D0 African Y-Chromosomal Haplogroup and Its Implications for the Expansion of Modern Humans out of Africa”. Genetics: genetics.302368.2019.
[9] Hammer MF, Karafet TM, Park H, Omoto K, Harihara S, Stoneking M, Horai S (2006). “Dual origins of the Japanese: common ground for hunter-gatherer and farmer Y chromosomes”. Journal of Human Genetics. 51 (1): 47–58
[10] Tajima A, Hayami M, Tokunaga K, Juji T, Matsuo M, Marzuki S, Omoto K, Horai S (2004). “Genetic origins of the Ainu inferred from combined DNA analyses of maternal and paternal lineages”. Journal of Human Genetics. 49 (4): 187–93
[11] Hua Zhong, Hong Shi, Xue-Bin Qi, Zi-Yuan Duan, Ping-Ping Tan, Li Jin, Bing Su, and Runlin Z. Ma (2011), “Extended Y Chromosome Investigation Suggests Postglacial Migrations of Modern Humans into East Asia via the Northern Route.” Mol. Biol. Evol.28(1):717–727.
[12] Shi H, Zhong H, Peng Y, Dong YL, Qi XB, Zhang F, Liu LF, Tan SJ, Ma RZ, Xiao CJ, Wells RS, Jin L, Su B (October 2008). “Y chromosome evidence of earliest modern human settlement in East Asia and multiple origins of Tibetan and Japanese populations”. BMC Biology. 6: 45.
[13] https://www.yfull.com/tree/E/
[14] https://www.yfull.com/tree/C/
[15] Fu; et al. (2016).”The genetic history of Ice Age Europe”. 10.1038/nature17993; Такође: Sikora; et al. (2017).”Ancient genomes show social and reproductive behavior of early Upper Paleolithic foragers”. 10.1126/science.aao1807.
[16] Nagle, N.; et al. (2015). “Antiquity and diversity of aboriginal Australian Y-chromosomes”. American Journal of Physical Anthropology. 159 (3): 367–81.
[17] ISOGG, 2015 “Y-DNA Haplogroup C and its Subclades – 2015”
[18] Malyarchuk B, Derenko M, Denisova G, Wozniak M, Grzybowski T, Dambueva I, Zakharov I. Phylogeography of the Y-chromosome haplogroup C in northern Eurasia. Ann Hum Genet. 2010 Nov;74(6):539-46. doi: 10.1111/j.1469-1809.2010.00601.x. Epub 2010 Aug 20. PMID: 20726964.
[19] https://www.yfull.com/tree/F/
[20] https://www.yfull.com/tree/G/
[21] Rootsi S, Myres NM, Lin AA, et al. (December 2012). “Distinguishing the co-ancestries of haplogroup G Y-chromosomes in the populations of Europe and the Caucasus”. Eur. J. Hum. Genet. 20 (12): 1275–82.
[22] Исто
[23] Mathieson; et al. (2015).”Genome-wide patterns of selection in 230 ancient Eurasians”. 10.1038/nature16152
[24] https://www.yfull.com/tree/H/
[25] Martínez-Cruz, B., Mendizabal, I., Harmant, C. et al. Origins, admixture and founder lineages in European Roma. Eur J Hum Genet 24, 937–943 (2016). https://doi.org/10.1038/ejhg.2015.201
[26] Mathieson; et al. (2015).”Genome-wide patterns of selection in 230 ancient Eurasians”. 10.1038/nature16152; Такође: Lazaridis; et al. (2016).”Genomic insights into the origin of farming in the ancient Near East”. 10.1038/nature19310; Lipson; et al. (2017).”Parallel palaeogenomic transects reveal complex genetic history of early European farmers”. 10.1038/nature24476.
[27] https://www.yfull.com/tree/I/
[28] Fu; et al. (2016).”The genetic history of Ice Age Europe”. 10.1038/nature17993
[29] https://www.yfull.com/tree/J/
[30] Chiaroni, Jacques; King, Roy J; Myres, Natalie M; Henn, Brenna M; Ducourneau, Axel; Mitchell, Michael J; Boetsch, Gilles; Sheikha, Issa; et al. (2010). “The emergence of Y-chromosome haplogroup J1e among Arabic-speaking populations”. European Journal of Human Genetics. 18 (3): 348–353
[31] Balanovsky, O.; Dibirova, K.; Dybo, A.; Mudrak, O.; Frolova, S.; Pocheshkhova, E.; Haber, M.; Platt, D.; et al. (2011). “Parallel evolution of genes and languages in the Caucasus region”. Molecular Biology and Evolution. 28 (10): 2905–20
[32] El-Sibai, Mirvat; Platt, Daniel E.; Haber, Marc; Xue, Yali; Youhanna, Sonia C.; Wells, R. Spencer; Izaabel, Hassan; Sanyoura, May F.; et al. (2009). “Geographical structure of the Y-chromosomal genetic landscape of the Levant: a coastal-inland contrast”. Annals of Human Genetics. 73 (Pt 6): 568–581; Такође: King, R. J.; Özcan, S. S.; Carter, T.; Kalfoğlu, E.; Atasoy, S.; Triantaphyllidis, C.; Kouvatsi, A.; Lin, A. A.; et al. (2008). “Differential Y-chromosome Anatolian influences on the Greek and Cretan Neolithic”. Annals of Human Genetics. 72 (Pt 2): 205–214.
[33] https://www.yfull.com/tree/K/
[34] Исто
[35] https://www.yfull.com/tree/L/
[36] Sengupta, S.; Zhivotovsky, L. A.; King, R.; Mehdi, S. Q.; Edmonds, C. A.; Chow, C-E. T.; Lin, A. A.; Mitra, M.; Sil, S. K.; Ramesh, A.; Usha Rani, M. V.; Thakur, C. M.; Cavalli-Sforza, L. L.; Majumder, P. P.; Underhill, P. A. (2006). “Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists”. The American Journal of Human Genetics. 78 (2): 202–21.
[37] Di Cristofaro, Julie; Pennarun, Erwan; Mazières, Stéphane; Myres, Natalie M.; Lin, Alice A.; Temori, Shah Aga; Metspalu, Mait; Metspalu, Ene; Witzel, Michael; King, Roy J.; Underhill, Peter A.; Villems, Richard; Chiaroni, Jacques (2013). “Afghan Hindu Kush: Where Eurasian Sub-Continent Gene Flows Converge”. PLOS ONE. 8 (10): e76748. Такође: Cordaux, Richard; Aunger, Robert; Bentley, Gillian; Nasidze, Ivane; Sirajuddin, S. M.; Stoneking, Mark (3 February 2004). “Independent origins of Indian caste and tribal paternal lineages”. Current Biology. 14 (3): 231–235; Lacau, Harlette; Gayden, Tenzin; Regueiro, Maria; Chennakrishnaiah, Shilpa; Bukhari, Areej; Underhill, Peter A; Garcia-Bertrand, Ralph L; Herrera, Rene J (18 April 2012). “Afghanistan from a Y-chromosome perspective”. European Journal of Human Genetics. 20 (10): 1063–1070.
[38] Vincenza Battaglia, Simona Fornarino, Nadia Al-Zahery, et al. (2009), “Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe.” European Journal of Human Genetics (2009) 17, 820–830; Такође: Oleg Balanovsky, Khadizhat Dibirova, Anna Dybo, et al. (2011), “Parallel Evolution of Genes and Languages in the Caucasus Region.” Molecular Biology and Evolution 2011 October ; 28(10): 2905–2920; Bayazit Yunusbayev, Mait Metspalu, Mari Järve, et al. (2012), “The Caucasus as an Asymmetric Semipermeable Barrier to Ancient Human Migrations.” Molecular Biology and Evolution 29(1):359–365.
[39] https://www.yfull.com/tree/T/
[40] Mendez FL, Karafet TM, Krahn T, Ostrer H, Soodyall H, Hammer MF (2011). “Increased resolution of Y chromosome haplogroup T defines relationships among populations of the Near East, Europe, and Africa”. Human Biology. 83 (1): 39–53.
[41] Lazaridis; et al. (2016).”Genomic insights into the origin of farming in the ancient Near East”. Nature, nature19310; Такође: Mathieson; et al. (2015).”Genome-wide patterns of selection in 230 ancient Eurasians”. Nature, volume528, 499–503.
[42] https://www.yfull.com/tree/M/
[43] Kayser, Manfred; Brauer, Silke; Weiss, Gunter; Schiefenhövel, Wulf; Underhill, Peter; Shen, Peidong; Oefner, Peter; Tommaseo-Ponzetta, Mila; Stoneking, Mark (2003). “Reduced Y-Chromosome, but Not Mitochondrial DNA, Diversity in Human Populations from West New Guinea”. The American Journal of Human Genetics. 72 (2): 281–302. Такође: Karafet, T. M.; Mendez, F. L.; Meilerman, M. B.; Underhill, P. A.; Zegura, S. L.; Hammer, M. F. (2008). “New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree”. Genome Research. 18 (5): 830–8.
[44] Nagle, N.; et al. (2015). “Antiquity and diversity of aboriginal Australian Y-chromosomes”. American Journal of Physical Anthropology. 159 (3): 367–81.
[45] https://www.yfull.com/tree/S/
[46] Kayser, Manfred; Brauer, Silke; Weiss, Gunter; Schiefenhövel, Wulf; Underhill, Peter; Shen, Peidong; Oefner, Peter; Tommaseo-Ponzetta, Mila; Stoneking, Mark (2003). “Reduced Y-Chromosome, but Not Mitochondrial DNA, Diversity in Human Populations from West New Guinea”. The American Journal of Human Genetics. 72 (2): 281–302.
[47] Scheinfeldt, L.; Friedlaender, F; Friedlaender, J; Latham, K; Koki, G; Karafet, T; Hammer, M; Lorenz, J (2006). “Unexpected NRY Chromosome Variation in Northern Island Melanesia”. Molecular Biology and Evolution. 23 (8): 1628–41. Такође: Cox, Murray P.; Mirazón Lahr, Marta (2006). “Y-chromosome diversity is inversely associated with language affiliation in paired Austronesian- and Papuan-speaking communities from Solomon Islands”. American Journal of Human Biology. 18 (1): 35–50.
[48] https://www.yfull.com/tree/P/
[49] Sikora; et al. (2019). “The population history of northeastern Siberia since the Pleistocene”. Nature 570, 182–188
[50] https://www.yfull.com/tree/Q/
[51] Jeffrey, T.; et al. (Jan 2002). “The Dual Origin and Siberian Affinities of Native American Y Chromosomes”. Am J Hum Genet. 70 (1): 192–206. Такође: Malyarchuk, Boris; et al. (2011). “Ancient links between Siberians and Native Americans revealed by subtyping the Y chromosome haplogroup Q1a”. Journal of Human Genetics. 56 (8): 583–588; Dulik; et al. (2012).” Mitochondrial DNA and Y Chromosome Variation Provides Evidence for a Recent Common Ancestry between Native Americans and Indigenous Altaians”. Am J Hum Genet., 90(2): 229–246.
[52] Zegura, S. L.; Karafet, TM; Zhivotovsky, LA; Hammer, MF (2004). “High-Resolution SNPs and Microsatellite Haplotypes Point to a Single, Recent Entry of Native American Y Chromosomes into the Americas” (PDF). Molecular Biology and Evolution. 21 (1): 164–75. Такође: Malhi, RS; Gonzalez-Oliver, A; Schroeder, KB; Kemp, BM; Greenberg, JA; Dobrowski, SZ; Smith, DG; Resendez, A; Karafet, T; Hammer, M; Zegura, S; Brovko, T (2008). “Distribution of Y chromosomes among native North Americans: a study of Athapaskan population history”. Am. J. Phys. Anthropol. 137 (4): 412–24.
[53] https://www.yfull.com/tree/R/
[54] Allentoft; et al. (2015). “Population genomics of Bronze Age Eurasia”. Nature, volume 522, 167–172. Такође: Mittnik; et al. (2018).”The genetic prehistory of the Baltic Sea region”. Nature Communications volume 9, Article number: 442.
[55] https://www.yfull.com/tree/R1b/
[56] Fu; et al. (2016). “The genetic history of Ice Age Europe”. 10.1038/nature17993
[57] Mathieson; et al. (2018). “The genomic history of southeastern Europe”. 10.1038/nature25778
[58] Wood, ET; Stover, DA; Ehret, C; Destro-Bisol, G; Spedini, G; Mcleod, H; Louie, L; Bamshad, M; et al. (2005). “Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes” (PDF). European Journal of Human Genetics. 13 (7): 867–76.
[59] D’Atanasio; et al. (2018). “The peopling of the last Green Sahara revealed by high-coverage resequencing of trans-Saharan patrilineages”. Genome Biol 19, 20
[60] https://www.yfull.com/tree/R2/
[61] Di Cristofaro, Julie; Pennarun, Erwan; Mazières, Stéphane; Myres, Natalie M.; Lin, Alice A.; Temori, Shah Aga; Metspalu, Mait; Metspalu, Ene; Witzel, Michael; King, Roy J.; Underhill, Peter A.; Villems, Richard; Chiaroni, Jacques (2013). “Afghan Hindu Kush: Where Eurasian Sub-Continent Gene Flows Converge”. PLOS ONE. 8 (10): e76748.
[62] https://www.yfull.com/tree/N/
[63] Rootsi, Siiri; Zhivotovsky, Lev A; Baldovič, Marian; Kayser, Manfred; Kutuev, Ildus A; Khusainova, Rita; Bermisheva, Marina A; Gubina, Marina; et al. (2006). “A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe”. European Journal of Human Genetics. 15 (2): 204–11.
[64] Ilumäe; et al. (2016). “Human Y Chromosome Haplogroup N: A Non-trivial Time-Resolved Phylogeography that Cuts across Language Families”. American Journal of Human Genetics. 99 (1): 163–73.
[65] Исто
[66] Српски ДНК пројекат; https://dnk.poreklo.rs/DNK-projekat/
[67] https://www.yfull.com/tree/O/
[68] Hammer, Michael F.; Karafet, Tatiana M.; Park, Hwayong; Omoto, Keiichi; Harihara, Shinji; Stoneking, Mark; Horai, Satoshi (2005). “Dual origins of the Japanese: Common ground for hunter-gatherer and farmer Y chromosomes”. Journal of Human Genetics. 51 (1): 47–58.
[69] Kim, SH; Kim, Ki-Cheol; Shin, Dong-Jik; Jin, Han-Jun; Kwak, Kyoung-Don; Han, Myun-Soo; Song, Joon-Myong; Kim, Won; Kim, Wook (2011). “High frequencies of Y-chromosome haplogroup O2b-SRY465 lineages in Korea: a genetic perspective on the peopling of Korea”. Investigative Genetics. 2 (10)
[70] Kumar; et al. (2007). “Y-chromosome evidence suggests a common paternal heritage of Austro-Asiatic populations”. BMC Evolutionary Biology 2007, 7:47; Такође: Reddy BM, Langstieh BT, Kumar V, Nagaraja T, Reddy ANS, et al (2007) Austro-Asiatic Tribes of Northeast India Provide Hitherto Missing Genetic Link between South and Southeast Asia. PLoS ONE 2(11): e1141.
[71] Kayser, M.; Choi, Y.; Van Oven, M.; Mona, S.; Brauer, S.; Trent, R. J.; Suarkia, D.; Schiefenhovel, W.; Stoneking, M. (2008). “The Impact of the Austronesian Expansion: Evidence from mtDNA and Y Chromosome Diversity in the Admiralty Islands of Melanesia”. Molecular Biology and Evolution. 25 (7): 1362–74; Такође: Karafet, T. M.; Hallmark, B.; Cox, M. P.; Sudoyo, H.; Downey, S.; Lansing, J. S.; Hammer, M. F. (2010). “Major East-West Division Underlies Y Chromosome Stratification across Indonesia”. Molecular Biology and Evolution. 27 (8): 1833–44.
[72] Shi, Hong; Dong, Yong-li; Wen, Bo; Xiao, Chun-Jie; Underhill, Peter A.; Shen, Pei-Dong; Chakraborty, Ranajit; Jin, Li; Su, Bing (2005). “Y-Chromosome Evidence of Southern Origin of the East Asian–Specific Haplogroup O3-M122”. The American Journal of Human Genetics. 77 (3): 408–19.
[73] https://www.yfull.com/tree/, приступљено 29.11.2025.
[74] https://www.familytreedna.com/public/y-dna-haplotree/, приступљено 29.11.2025.
[75] Видети: https://dnk.poreklo.rs/DNK-projekat/.








-270x270.jpg)


















Коментари (0)