



Srpski DNK projekat je pokrenuo objavljivanje serije priloga u kojima čitaoce upoznaje sa osnovama genetičke genealogije, discipline koja poslednjih godina nailazi na sve veće interesovanje. Serijal je nastao kao dopunjena i prilagođena verzija članka „Genetika u službi rodoslovlja“ (V.Mihić, M. Rajevac, S. Jerković), objavljenog u knjizi „Genetičko poreklo Srba Stare Hercegovine“ (2021). Ovakva vrsta sadržaja do sada nije bila dostupna na srpskom jeziku u formi naučno-popularne građe onima koji su želeli da se upoznaju sa osnovama ove nove naučne oblasti. Zato smo pripremili „Malu školu genetičke genealogije“ kroz koju ćemo dati jasna objašnjenja i primere koji približavaju pojmove i metode savremene genetike, kao i njihovu primenu u istraživanju porekla porodica, rodova i naroda.
U prvom nastavku serijala predstavili smo osnovne pojmove genetike: šta su i kako funkcionišu geni i DNK, šta su hromozomi i kako se odvija reprodukcija i nasleđivanje. Bilo je reči i o ljudskom genomu, genetičkim mutacijama, kao i o STR i SNP markerima koji čine osnovu savremenih genetičkih testova.
Drugi nastavak koji sledi donosi objašnjenje vrsta i namene genetičkih testova, kao i pojam haplogrupa. Čitaoci će se upoznati sa filogenetičkim stablima i metodama određivanja pripadnosti Y‑DNA haplogrupi, uz primer kako se rezultati tumače i povezuju sa rodoslovnim istraživanjima.
GENETIČKI TESTOVI
Genetički testovi koji se rade u genealoške svrhe baziraju se na poređenju sličnosti, odnosno pronalaženju malih razlika[1] u strukturi genoma dve ili više osoba. Svaki genetički test sprovodi se u dva osnovna koraka – faza utvrđivanja strukture DNK i faza analize strukture DNK:
I. Utvrđivanje strukture DNK ovde podrazumeva različite laboratorijske procese i metode kojima se tačno određuje tip i redosled nukleobaza u nukleotidnim lancima DNK uzorka. Tehnikom sekvenciranja može se utvrditi redosled nukleobaza u dužim ili kraćim segmentima DNK, pojedinačnim hromozomima, mitohondrijskoj DNK ili u celom genomu, dok se drugim metodama, poput genotipiziranja, mogu očitavati genetički markeri na ciljanim lokusima.[2]
II. Analiza ustanovljene strukture DNK je faza genetičkog testa u kojoj se na osnovu različitih algoritama i statističkih modela prethodno utvrđena struktura DNK analizira i tumači u cilju izvođenja zaključaka koji se dalje koriste za različite namene – u forenzici, genealogiji, medicini i drugim naučnim i stručnim oblastima.
I. Struktura DNK može se utvrđivati za različite delove ljudskog genoma:
1. Očitavanje predefinisanih lokusa DNK
Ovim metodama određuju se nukleobaze na tačno određenim pozicijama u DNK lancu čiji je sadržaj od interesa ili se utvrđuje broj STR ponovaka na izabranim lokusima. Na taj način se očitavaju vrednosti na ciljanim pozicijama koje nazivamo SNP i STR markerima. SNP marker predstavlja konkretnu varijaciju na određenom lokusu, dok STR marker označava broj ponovaka karakteristične sekvence na nekom lokusu. Pored provere pojedinačnih genetičkih markera u komercijalnoj ponudi su i panel-testovi kao i čip-testovi kojima se može određivati veći broj SNP ili STR markera. U genealoške svrhe proveravaju se najčešće SNP i STR markeri na Y hromozomu, autozomni SNP markeri[3], a ređe i SNP markeri na mitohondrijskoj DNK.
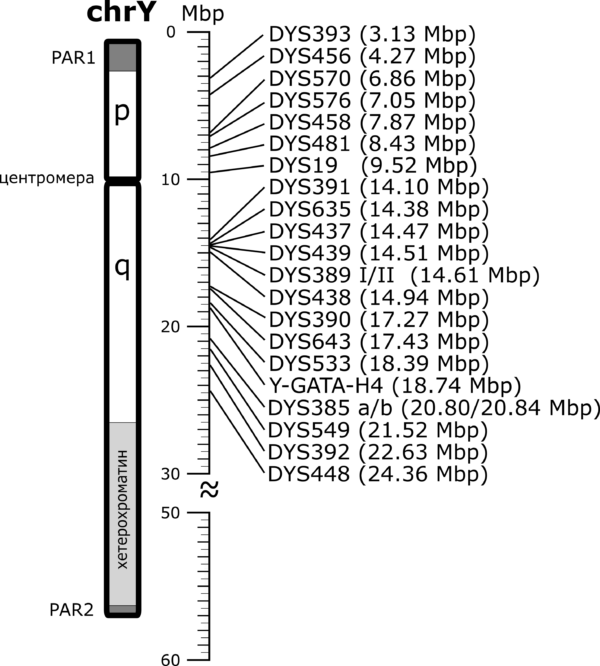
Pozicije YSTR markera iz PowerPlex Y23 test panela u odnosu na Y hromozom, prema GRCh37 referentnom modelu ljudskog genoma.[4]
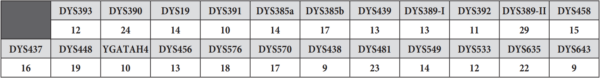
Testovi kojima se određuju vrednosti STR markera na Y hromozomu nazivaju se Y-STR testovi. Iako je moguća provera vrednosti i za pojedinačne STR markere, laboratorije i kompanije koje se bave genetičkim testiranjem nude standardizovane panel-testove kojima se određuje različit broj i skup Y-STR markera. Panelima se obično određuje 12, 17, 23, 37, 67 ili 111 Y-STR markera. U laboratorijama u Srbiji trenutno je dostupan takozvani PowerPlex Y23 panel[5], kojim se utvrđuje vrednost 23 YSTR markera, kao i Investigator Argus Y-28 QS Kit[6], kojim se analizira 28 Y-STR markera, a do pre petnaesetak godina radio se uglavnom AmpFLSTR Yfiler panel[7], koji obuhvata skup od 17 markera. Rezultati ovakvih testova predstavljaju niz vrednosti markera na lokusima na kojima se proverava broj ponovaka. Ovaj niz predstavlja svojevrstan DNK potpis osobe čiji se hromozom analizira i naziva se haplotip. Logično, u zavisnosti od odabranog test-panela, haplotip može biti određen na manjem ili većem broju YSTR markera. Na primer, jedan haplotip određen za 23 YSTR markera čije se vrednosti utvrđuju PowerPlex Y23 komercijalnim test-panelom izgleda ovako:
2. Sekvenciranje dužih segmenta genoma
Kod ove vrste testova DNK sekvenciranjem se određuje struktura jednog dela genoma. U komercijalnoj ponudi su testovi kojima se sekvencira Y hromozom (Y-DNA) ili mitohondrijska DNK (mtDNA). U praksi se retko doseže stoprocentna pokrivenost ciljanog DNK segmenta, a često za time i ne postoji potreba ukoliko se testiranje radi u genealoške svrhe. Y hromozom je sastavljen od približno 62,5 miliona baznih parova[8], od kojih je trenutno manje od 40% od genealoškog značaja[9], pa neki komercijalno raspoloživi testovi sekvenciraju samo taj segment DNK lanca Y hromozoma. U ovakvu vrstu testova spada poznati Big Y-700 test kompanije FamilyTreeDNA [10].
3. Sekvenciranje celog genoma
Cena testova kojima se utvrđuje DNK struktura kompletnog ljudskog genoma (eng. Whole Genome Sequencing), to jest vrši sekvenciranje svih 23 para hromozoma i mitohondrijske DNK postaje sve pristupačnija. Neke kompanije garantuju stoprocentnu pokrivenost genoma testom, mada se u praksi dešava da ona bude nešto manja.
II. Prethodno utvrđena struktura DNK može se dalje analizirati i obrađivati u genealoške svrhe:
1. Genetička analiza autozomnih hromozoma
Ova vrsta analize ispituje genetičku sličnost dve ili više osoba na osnovu 22 para homolognih autozomnih hromozoma. Srodnost testiranih pojedinaca može se porediti po svim njihovim predačkim naslednim linijama. Zbog prethodno objašnjene specifičnosti nasleđivanja, koja se ogleda u činjenici da svaka osoba na svoje potomstvo prenosi samo 50% svog slučajno odabranog DNK materijala sadržanog u hromozomima, predačke linije se ovom metodom analize obično mogu relativno pouzdano porediti do pet generacija u prošlost[11].
Analiza autozomnih hromozoma može se koristiti za utvrđivanje geografskog porekla predaka i veoma je popularna među stanovnicima zemalja Novog sveta (Amerike, Australija, Novi Zeland). Takođe je često koriste osobe koje tragaju za svojim bližim biološkim srodnicima.
Postoje različite tehnike analize autozomnih hromozoma. Jedna od glavnih metoda koje se u tu svrhu koriste jeste pronalaženje takozvanih IBD segmenata (IBD: eng. Identical by descent – identičan po poreklu), a primenljiva je za poređenja srodnosti osoba koje su nasledile isti autozomni hromozom od zajedničkog pretka. Princip ove metode zasniva se na pronalaženju podudarnih delova između odgovarajućih autozomnih hromozoma pojedinaca čija se genetička srodnost proverava. Ovi delovi se nazivaju IBD segmentima. Tokom gametogeneze, pojava genetičke rekombinacije na parovima homolognih hromozoma dovodi do razmene delova između njihovih DNK lanaca. To uzrokuje svojevrsnu fragmentaciju originalne strukture hromozoma koji se dalje prenosi na potomstvo. Sa svakom novom generacijom na koju se prenosi određeni autozomni hromozom, on se dodatno fragmentiše genetičkom rekombinacijom. Posledično, što je veća srodnička udaljenost između dve osobe koje nose zajednički predački autozomni hromozom, biće sve kraći IBD segmenti koje one na tom hromozomu dele, pa se može uspostaviti korelacija između dužine IBD segmenata i broja generacija do zajedničkog pretka osoba koje su u srodstvu.
Druga metoda analize autozomnih hromozoma zasniva se na poređenju SNP markera, odnosno varijacija na tačno određenim SNP lokusima. U komercijalnim testovima, poput onih koji nude 23andMe, MyHeritage, AncestryDNA ili FTDNA Family Finder, analizira se nekoliko stotina hiljada do preko 700.000 SNP markera[12] raspoređenih po svim autozomnim hromozomima. Ovakvi testovi omogućavaju da se utvrdi genetička srodnost između osoba, ali i da se proceni geografsko poreklo predaka kroz poređenje sa referentnim populacijama. Laboratorije koriste biočipove dizajnirane za masovno očitavanje SNP markera (genotipizacija), što omogućava brzu i pouzdanu analizu velikog broja genetičkih varijacija.
2. Genetička analiza mitohondrijske DNK
Analiza mitohondrijske DNK omogućava poređenje srodnosti osoba po matrilinearnom genetičkom nasleđu. Mitohondrijska DNK se uvek prenosi gotovo neizmenjena isključivo sa majke na dete, pa je putem nje moguće pratiti genetičku srodnost pojedinaca do daleke prošlosti. Problemi sa ovom vrstom analize su kako biološke tako i sociološke prirode. Biološki problem leži u učestalosti pojavljivanja mutacija na mitohondrijskoj DNK, koje nastaju mnogo ređe nego što je to slučaj sa mutacijama na Y hromozomima.[13] Iako se na prvi pogled ovo čini dobrom karakteristikom, nedostatak mutacija dovodi do prevelike uniformnosti mitohondrijske DNK, odnosno do slabog grananja mtDNA filogenetičkog stabla, zbog čega se teže dolazi do pouzdanih zaključaka o stvarnoj rodbinskoj bliskosti testiranih pojedinaca.
Sociološki problem mtDNA analize je to što su ljudska društva od pradavnih vremena zasnovana na patrijarhatu, te da se posledično u istorijskim izvorima uglavnom mogu pronalaziti podaci za patrilinearne nasledne linije, dok ostale retko sežu dalje od nekoliko generacija. U kombinaciji sa slabom diversifikacijom mtDNA genetičkog stabla, nasleđena sociološka ograničenja čine mtDNA analizu manje upotrebljivom u genealoške svrhe.
3. Genetička analiza Y hromozoma
Ova vrsta analize DNK zasniva se na poređenju Y hromozoma između testiranih pojedinaca. Kako ovaj hromozom ne poseduju žene, pomoću Y-DNA analize može se pratiti samo patrilinearna rodbinska veza, to jest genetičko nasleđe koje se prenosi isključivo sa oca na sina. Za tradicionalno rodoslovlje koje prati muške nasledne linije ova vrsta analiza je od najvećeg značaja. Postoje dve vrste analiza Y hromozoma, od kojih se jedna zasniva na poređenju STR markera, dok se druga zasniva na pronalaženju SNP varijacija.
FILOGENETIČKA STABLA I HAPLOGRUPE
Jednom stečene SNP varijacije na Y hromozomu prenose se sa oca na sina, čime se u svakom čoveku akumuliraju promene koje su nastale na ovom hromozomu kod svih njegovih predaka po muškoj naslednoj liniji. Pažljivim poređenjem prisutnih SNP varijacija kod testiranih pojedinaca i primenom heurističkih i statističkih metoda naučnici su uspeli da formiraju Y-DNA filogenetičko stablo ljudskog roda. Ovo stablo predstavlja svojevrsni rodoslov muške populacije čovečanstva, a njegovo grananje određeno je prisutnim SNP varijacijama. U korenu ovog stabla nalazi se „Y-hromozomski Adam“, odnosno rekonstruisani Y hromozom muškarca koji je pretpostavljeni najstariji patrilinearni predak svih dosad testiranih muškaraca. On je prema trenutnim proračunima živeo pre 237.000 do 581,000 godina.[14] Važno je razumeti da to nije bio jedini muškarac vremena u kojem je živeo, ali se samo njegova muška loza nastavila bez prekida sve do danas. S obzirom da se radi o rekonstruisanom modelu Y hromozoma našeg najstarijeg muškog pretka, „titula“ Y-hromozomskog Adama se sa novim otkrićima još starijih grana Y-DNA filogenetičkog stabla može pomerati još dalje u prošlost.[15]
Sa rastom broja testiranih pojedinaca i otkrivanjem novih SNP varijacija Y-DNA filogenetičko stablo se stalno dalje grana i unapređuje.[16] Statističkim metodama se može odrediti verovatnoća pojavljivanja SNP mutacija u svakoj novoj generaciji, pa je određenim proračunima moguće proceniti vreme formiranja pojedinačnih grana filogenetičkog stabla.
Za sve pojedince kod kojih je prisutna određena SNP varijacija koja formira neku granu na Y-DNA filogenetičkom stablu kažemo da pripadaju Y-DNA haplogrupi koja je određena tom varijacijom. Tako na primer, svi muškarci koji poseduju SNP varijaciju I-M170 pripadaju haplogrupi I-M170, koja se još naziva i haplogrupa I i formirana je pre oko 43.000 godina. Ova grana filogenetičkog stabla dalje se grana na sve mlađe podgrane, određene mlađim SNP mutacijama, koje definišu i mlađe haplogrupe. Svaka mlađa grana u sebi nosi sve SNP varijacije prisutne u starijim granama iz kojih proističe. Tako na primer, pripadnici grana I1 i I2 nose SNP varijaciju I-M170, koja je karakteristična za sve pripadnike haplogrupe I iz koje ove grane proizilaze, ali takođe poseduju i svoje karakteristične SNP varijacije, specifične samo za granu I1, odnosno I2.
Dogovorom je usvojena podela stabla na dvadesetak glavnih haplogrupa, koje se označavaju velikim slovima engleskog pisma, a koje se dalje granaju na brojne podgrane.
Poput Y-DNA filogenetičkog stabla, postoji i mtDNA filogenetičko stablo, koje se grana po SNP varijacijama otkrivenim u mitohondrijskoj DNK i predstavlja svojevrsno matrilinearno rodoslovno stablo ljudskog roda. Ekvivalentno, postoje takođe i mtDNA haplogrupe. Slično „Y‑hromozomskom Adamu“, u genetičkoj genealogiji postoji i pojam „mitohondrijske Eve“. On označava najstarijeg zajedničkog matrilinearnog pretka svih današnjih ljudi, odnosno ženu čija se mitohondrijska DNK prenela na sve žive osobe putem majčine nasledne linije. Kao i u slučaju Y‑hromozomskog Adama, radi se o statističkom modelu zasnovanom na analizi SNP varijacija u mitohondrijskoj DNK. Mitohondrijska Eva nije bila jedina žena svog vremena, već jedina čija je matrilinearna linija opstala do danas bez prekida. Na osnovu molekularnog časovnika procenjuje se da je mitohondrijska Eva živela pre otprilike 150.000 do 200.000 godina u Africi, što se poklapa sa arheogenetičkim nalazima i modelima o afričkom poreklu savremenog čoveka.[17]
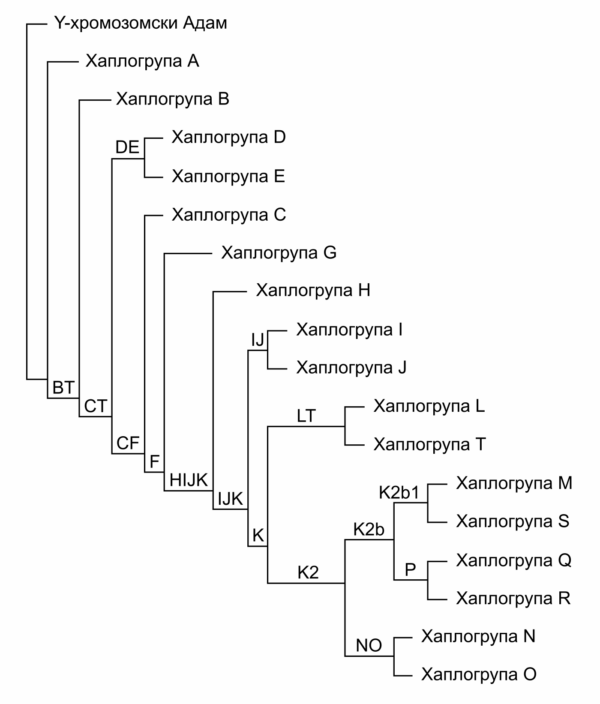 Pojednostavljeno filogenetičko stablo Y-DNA haplogrupa. [18]
Pojednostavljeno filogenetičko stablo Y-DNA haplogrupa. [18]
ODREĐIVANJE PRIPADNOSTI Y-DNA HAPLOGRUPI
Za pouzdano određivanje pripadnosti nekoj haplogrupi potrebno je uraditi Y-SNP test kojim se potvrđuje prisustvo ciljane SNP varijacije. S obzirom na to da postoji veoma veliki broj mogućih SNP varijacija čije bi se prisustvo moglo proveravati, ovakvi testovi, kojima se ispituju sve SNP varijacije na hromozomu, prilično su skupi. Oni spadaju u pominjane tipove testova kojima se sekvencira određeni segment genoma ili ceo genom. Moguće je i testiranje na pojedinačne SNP varijacije, ali ono predstavlja „traženje igle u plastu sena“, što opet može rezultovati velikim troškovima. Da bi se potraga za prisutnim SNP varijacijama suzila i imala prihvatljivu cenu, koristi se druga vrsta Y-DNA genetičkog testa – Y-STR test. Ideja se zasniva na pretpostavci da je pojava Y-SNP mutacija praćena odgovarajućim Y-STR mutacijama, te da pripadnici određene haplogrupe poseduju slične Y-STR haplotipove. Empirijskom analizom raspoloživih rezultata testiranja ovakva pretpostavka pokazala se tačnom. Svakoj haplogrupi odgovara određeni, manje ili više karakterističan haplotip, čiji se Y-STR markeri javljaju kod najvećeg broja pripadnika te haplogrupe. Ovakav haplotip naziva se modalni haplotip haplogrupe, a za markere koji ga formiraju kaže se da imaju modalnu vrednost.
Određivanjem Y-STR haplotipa testirane osobe može se s velikom verovatnoćom utvrditi haplogrupa kojoj ta osoba pripada, čime se značajno sužava broj SNP markera koje je potrebno dodatno proveravati u cilju otkrivanja najmlađe grane filogenetičkog stabla kojoj pripada testirani pojedinac. Treba ipak naglasiti da se utvrđivanje pripadnosti haplogrupi na osnovu Y-STR testa zasniva na principima teorije verovatnoće i matematičke statistike. Tu nam u pomoć dolaze računarski programi prediktori, koji na osnovu haplotipa određuju najverovatniju pripadnost haplogrupi. Što je veći broj utvrđenih YSTR markera u haplotipu, to se haplogrupa može utvrditi sa većom preciznošću i otkriti njena najmlađa grana. Ipak, za apsolutnu potvrdu pripadnosti nekoj haplogrupi neophodno je uraditi SNP test.
Slično kao za haplogrupu, modalni haplotip može se odrediti i za bilo koju grupu testiranih pojedinaca koji pripadaju istoj grani filogenetičkog stabla. Što su razlike na YSTR markerima između testiranih pojedinaca manje, to su veći izgledi da oni dele zajedničkog muškog pretka u bližoj prošlosti. Postoje algoritmi kojima se na osnovu sličnosti haplotipova može manje ili više pouzdano proceniti vreme do najmlađeg zajedničkog pretka dve osobe (engleski Time to most recent common ancestor, ili skraćeno TMRCA). Ove procene treba ipak uzimati s dozom rezerve, s obzirom na to da predstavljaju samo najverovatniji vremenski raspon, koji u stvarnosti može ponekad i značajno da varira.
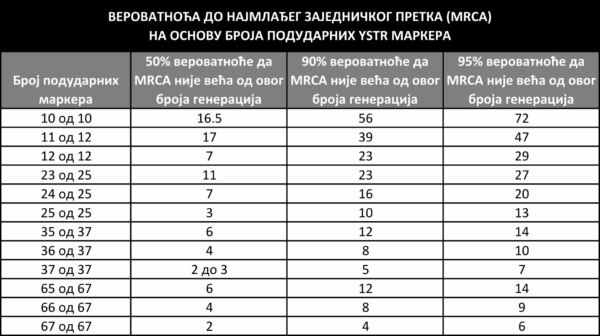
Jedna procena verovatnoće da osobe čiji se haplotipovi podudaraju
ili su bliski dele zajedničkog pretka u zadatom broju generacija. [18.2]
Verovatnoća pojave mutacije na YSTR markerima podleže zakonima koji se javljaju kod svih slučajnih prirodnih procesa: što se vrednost YSTR markera više razlikuje od modalne za posmatranu haplogrupu, to je manja verovatnoća da se ona pojavi. Takođe, što je određeni YSTR marker skloniji bržim promenama, to je (jezikom statistike rečeno) njegovo srednje apsolutno odstupanje od modalne vrednosti veće. Drugim rečima, kod haplotipova pripadnika određene haplogrupe, na brzo promenljivim markerima će se češće javljati vrednosti različite od modalne. Može se reći da vrednosti markera koje se retko pojavljuju predstavljaju osobenost haplotipova u kojima se javljaju, pa se za njih kaže da predstavljaju karakteristične vrednosti markera.
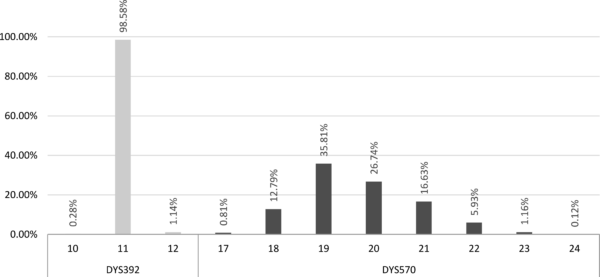
Učestalost pojavljivanja različitih vrednosti
sporo promenljivog markera DYS392 i brzo promenljivog DYS570 kod haplogrupe E-V13
Na slici je prikazan primer raspodele učestalosti pojavljivanja vrednosti markera DYS392 i DYS570 na jednom uzorku haplogrupe E-V13.[19] Kao što se na grafikonu vidi, za sporo promenljivi marker DYS392 modalna vrednost je 11 i javlja se u više od 98% slučajeva, dok su vrednosti 10 i 12 javljaju veoma retko i mogu se smatrati karakterističnim. Kod drugog markera, DYS570, modalna vrednost 19 javlja se kod tek nešto više od 35% testiranih pojedinaca. Kod ovog markera se vrednost 20 ne može smatrati karakterističnom, iako odstupa od modalne, s obzirom na to da se prilično često pojavljuje. Zbog velike varijacije markera DYS570 čak se i vrednosti 18 i 21 mogu smatrati tek nešto ređim, dok se jedino za vrednosti 17, 22, 23 i 24 može reći da su karakteristične.[20]
Sličnost haplotipova testiranih pojedinaca kod kojih se javljaju karakteristični markeri povećava verovatnoću da pripadaju istoj haplogrupi, to jest da među njima postoji bliža genetička veza, pod uslovom da podudarnost postoji na karakterističnim markerima. Važi i suprotno: o stvarnoj bliskosti pojedinaca koji poseduju modalne haplotipove, ili one koji ne poseduju karakteristične markere, nezahvalno je izvoditi zaključke bez dodatnih SNP testova.
Za grupu haplotipova koja poseduje karakteristične markere, koji je jasno izdvajaju iz osnovne haplogrupe kojoj pripadaju, kaže se da je dobro profilisana. Dobro profilisan skup haplotipova po pravilu pripada mlađoj grani osnovne haplogrupe, koju karakterišu i odgovarajuće SNP varijacije. Na primer, ukoliko haplotipovi grupe testiranih pojedinaca koji pripadaju haplogrupi I2-Z17855 poseduju karakteristične markere DYS385ab=14-14 i DYS448=21, oni po pravilu pripadaju mlađoj grani I2-PH3414. Ova grana je karakteristična za hercegovački rod Mirilovića, te se za sve haplotipove koji su bliski modalnom haplotipu haplogrupe I2-Z17855, a poseduju pomenute karakteristične YSTR markere, može osnovano pretpostaviti da pripadaju rodu Mirilovića. Sledeći ovu metodologiju moguće je izvoditi manje ili više pouzdane zaključke o pripadnosti haplotipova odgovarajućim granama filogenetičkog stabla, to jest genetičkim rodovskim grupama.
♦ ♦ ♦ ♦ ♦
U trećem nastavku daćemo pregled osnovnih Y‑DNA haplogrupa u svetu. Biće reči o njihovoj starosti, geografskoj rasprostranjenosti i značaju u istorijskom kontekstu, kao i o haplogrupama zastupljenim kod Srba.
[1] Ne treba zaboraviti da je 99.9% strukture genoma kod svih ljudi identično.
[2] Primenom savremenih tehnologija i naučnih saznanja proces sekvenciranja DNK vremenom se usavršavao i razvijao, što je praćeno i drastičnim padom cene sekvenciranja ljudskog genoma u 21. veku. S neverovatnih 100 miliona dolara, koliko su iznosili troškovi sekvenciranja jednog ljudskog genoma 2001. godine, današnja cena pala je na manje od 500 dolara (Videti: DNA Sequencing Costs: Data, National Human Genome Research Institute, 2023, https://www.genome.gov/about-genomics/fact-sheets/DNA-Sequencing-Costs-Data, pristupljeno 29.11.2025).
[3] Ovo se odnosi na testove kojima se ispituju autozomni hromozomi, poput 23andMe, MyHeritage FamilyFinder i sličnih. Ovi testovi se oslanjaju na metodu koja se naziva genotipizacija, a kojom se pomoću posebno dizajniranih biočipova istovremeno očitava po nekoliko stotina hiljada odabranih SNP markera.
[4] Izvor: Becky Hill, John Butler, and Mike Coble, Variability of Y-STR Marker Sets in the NIST 1036 U.S. Population Samples, National Institute of Standards and Technology, Bode Technology 1st Annual Advanced DNA Technology, Mid-Atlantic Workshop, Charlottesville, VA, USA, September 19 & 20, 2012.
[5] https://worldwide.promega.com/products/forensic-dna-analysis-ce/str-amplification/powerplex-y23-system/?catNum=DC2305, pristupljeno 29.11.2025.
[6] https://www.qiagen.com/us/products/human-id-and-forensics/investigator-solutions/investigator-argus-y-28-qs-kit, pristupljeno 29.11.2025.
[7] https://www.thermofisher.com/order/catalog/product/4359513, pristupljeno 29.11.2025.
[8] Rhie, A., Nurk, S., Cechova, M. et al. The complete sequence of a human Y chromosome. Nature 621, 344–354 (2023). https://doi.org/10.1038/s41586-023-06457-y
[9] S obzirom da je 2023. godine po prvi put u potpunosti sekvenciran i poslednji deo genoma koji do tada nije bio dekodiran, a koji je pripadao upravo Y hromozomu, u budućnosti će se za potrebe genetičke genealogije svakako analizirati veći deo Y hromozoma.
[10] Big Y-700: The Forefront Of Y Chromosome Testing, 7. June 2019, https://blog.familytreedna.com/human-y-chromosome-testing-milestones/. Takođe: Caleb Davis, Michael Sager, Göran Runfeldt, Elliott Greenspan, Arjan Bormans, Bennett Greenspan, and Connie Bormans, Big Y-700 White Paper. https://blog.familytreedna.com/wp-content/uploads/2019/03/big-y-700-white-paper_compressed.pdf (pristupljeno 29.11.2025).
[11] Autosomal DNA, https://isogg.org/wiki/Autosomal_DNA (pristupljeno 29.11.2025)
[12] https://support.ancestry.com/s/article/What-are-DNA-Markers?language=en_US, (pristupljeno 29.11.2025).
[13] Bert Ely, How do researchers trace mitochondrial DNA over centuries?, Scientific American, 2006, https://www.scientificamerican.com/article/how-do-researchers-trace/ (pristupljeno 29.11.2025).
[14] Videti: https://isogg.org/wiki/Y-chromosomal_Adam
[15] Godine 2012. u Kamerunu je otkriveno žarište do tada nepoznate prastare Y haplogrupe koja je dobila oznaku A00. Novootkrivena haplogrupa je pomerila vreme u kojem je živeo Y-hromozomski Adam desetinama hiljada godina u prošlost, čak za više od 100.000 godina. Videti: Mendez, Fernando L. et al. An African American Paternal Lineage Adds an Extremely Ancient Root to the Human Y Chromosome Phylogenetic Tree, The American Journal of Human Genetics, Volume 92, Issue 3, 454 – 459, 2013, https://doi.org/10.1016/j.ajhg.2013.02.002
[16] Kako genetičke testove i analize sprovode različite organizacije i laboratorije, nažalost, ne postoji jedinstveno standardizovano Y-DNA filogenetičko stablo. Iako je osnovna struktura ovih stabala u svim slučajevima ista, postoje razlike u vidu prisustva ili odsustva nekih mlađih grana. Jedan od zadataka organizacije koja nosi naziv „Međunarodno udruženje za genetičku genealogiju“ (International Society of Genetic Genealogy, https://isogg.org/) je da radi na standardizaciji strukture i nomenklature Y-DNA filogenetičkog stabla.
[17] ISOGG Wiki – Mitochondrial Eve, https://isogg.org/wiki/Mitochondrial_Eve (pristupljeno 29.11.2025).
[18] Izvor: https://www.yfull.com/tree/ (pristupljeno 29.11.2025).
[18.2] Izvor: https://www.myfamilydnatest.com/ftdna-y-37-vs-y-67-vs-y-111-vs-y-500-tests/ (pristupljeno 8.8.2022).
[19] Reč je o uzorku sačinjenom od haplotipova haplogrupe E-V13 koji se nalaze u bazi Srpskog DNK projekta.
[20] Osobina karakterističnosti markera ne zavisi, dakle, samo od učestalosti pojavljivanja njegove vrednosti unutar posmatranog skupa haplotipova, već i od stepena odstupanja te učestalosti u odnosu na učestalost pojavljivanja njegove modalne vrednosti unutar iste grupe haplotipova.








-270x270.jpg)


















Komentari (0)