



Српски ДНК пројекат је покренуо објављивање серије прилога у којима читаоце упознаје са основама генетичке генеалогије, дисциплине која последњих година наилази на све веће интересовање. Серијал је настао као допуњена и прилагођена верзија чланка „Генетика у служби родословља“ (В.Михић, М. Рајевац, С. Јерковић), објављеног у књизи „Генетичко порекло Срба Старе Херцеговине“ (2021). Оваква врста садржаја до сада није била доступна на српском језику у форми научно-популарне грађе онима који су желели да се упознају са основама ове нове научне области. Зато смо припремили „Малу школу генетичке генеалогије“ кроз коју ћемо дати јасна објашњења и примере који приближавају појмове и методе савремене генетике, као и њихову примену у истраживању порекла породица, родова и народа.
У првом наставку серијала представили смо основне појмове генетике: шта су и како функционишу гени и ДНК, шта су хромозоми и како се одвија репродукција и наслеђивање. Било је речи и о људском геному, генетичким мутацијама, као и о STR и SNP маркерима који чине основу савремених генетичких тестова.
Други наставак који следи доноси објашњење врста и намене генетичких тестова, као и појам хаплогрупа. Читаоци ће се упознати са филогенетичким стаблима и методама одређивања припадности Y‑DNA хаплогрупи, уз пример како се резултати тумаче и повезују са родословним истраживањима.
ГЕНЕТИЧКИ ТЕСТОВИ
Генетички тестови који се раде у генеалошке сврхе базирају се на поређењу сличности, односно проналажењу малих разлика[1] у структури генома две или више особа. Сваки генетички тест спроводи се у два основна корака – фаза утврђивања структуре ДНК и фаза анализе структуре ДНК:
I. Утврђивање структуре ДНК овде подразумева различите лабораторијске процесе и методе којима се тачно одређује тип и редослед нуклеобаза у нуклеотидним ланцима ДНК узорка. Техником секвенцирања може се утврдити редослед нуклеобаза у дужим или краћим сегментима ДНК, појединачним хромозомима, митохондријској ДНК или у целом геному, док се другим методама, попут генотипизирања, могу очитавати генетички маркери на циљаним локусима.[2]
II. Анализа установљене структуре ДНК је фаза генетичког теста у којој се на основу различитих алгоритама и статистичких модела претходно утврђена структура ДНК анализира и тумачи у циљу извођења закључака који се даље користе за различите намене – у форензици, генеалогији, медицини и другим научним и стручним областима.
I. Структура ДНК може се утврђивати за различите делове људског генома:
1. Очитавање предефинисаних локуса ДНК
Овим методама одређују се нуклеобазе на тачно одређеним позицијама у ДНК ланцу чији је садржај од интереса или се утврђује број STR поновака на изабраним локусима. На тај начин се очитавају вредности на циљаним позицијама које називамо SNP и STR маркерима. SNP маркер представља конкретну варијацију на одређеном локусу, док STR маркер означава број поновака карактеристичне секвенце на неком локусу. Поред провере појединачних генетичких маркера у комерцијалној понуди су и панел-тестови као и чип-тестови којима се може одређивати већи број SNP или STR маркера. У генеалошке сврхе проверавају се најчешће SNP и STR маркери на Y хромозому, аутозомни SNP маркери[3], а ређе и SNP маркери на митохондријској ДНК.
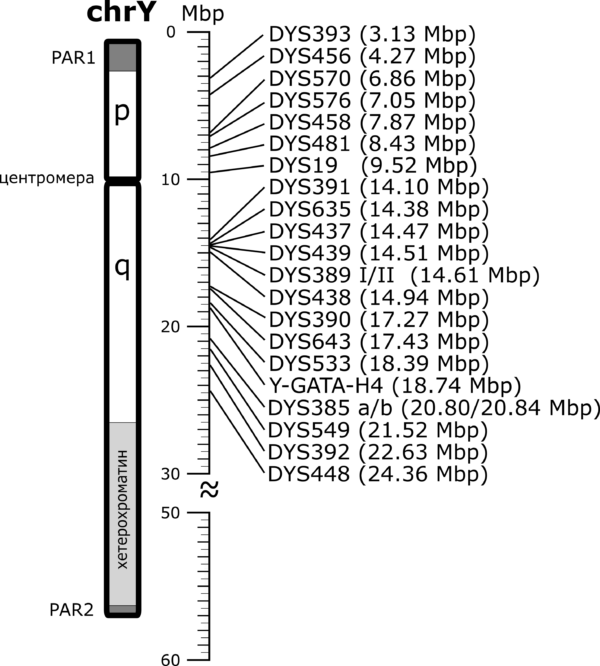
Позиције YSTR маркера из PowerPlex Y23 тест панела у односу на Y хромозом, према GRCh37 референтном моделу људског генома.[4]
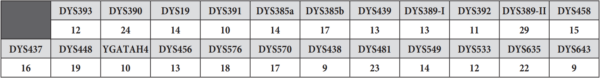
Тестови којима се одређују вредности STR маркера на Y хромозому називају се Y-STR тестови. Иако је могућа провера вредности и за појединачне STR маркере, лабораторије и компаније које се баве генетичким тестирањем нуде стандардизоване панел-тестове којима се одређује различит број и скуп Y-STR маркера. Панелима се обично одређује 12, 17, 23, 37, 67 или 111 Y-STR маркера. У лабораторијама у Србији тренутно је доступан такозвани PowerPlex Y23 панел[5], којим се утврђује вредност 23 YSTR маркера, као и Investigator Argus Y-28 QS Kit[6], којим се анализира 28 Y-STR маркера, а до пре петнаесетак година радио се углавном AmpFLSTR Yfiler панел[7], који обухвата скуп од 17 маркера. Резултати оваквих тестова представљају низ вредности маркера на локусима на којима се проверава број поновака. Овај низ представља својеврстан ДНК потпис особе чији се хромозом анализира и назива се хаплотип. Логично, у зависности од одабраног тест-панела, хаплотип може бити одређен на мањем или већем броју YSTR маркера. На пример, један хаплотип одређен за 23 YSTR маркера чије се вредности утврђују PowerPlex Y23 комерцијалним тест-панелом изгледа овако:
2. Секвенцирање дужих сегмента генома
Код ове врсте тестова ДНК секвенцирањем се одређује структура једног дела генома. У комерцијалној понуди су тестови којима се секвенцира Y хромозом (Y-DNA) или митохондријска ДНК (mtDNA). У пракси се ретко досеже стопроцентна покривеност циљаног ДНК сегмента, а често за тиме и не постоји потреба уколико се тестирање ради у генеалошке сврхе. Y хромозом је састављен од приближно 62,5 милиона базних парова[8], од којих је тренутно мање од 40% од генеалошког значаја[9], па неки комерцијално расположиви тестови секвенцирају само тај сегмент ДНК ланца Y хромозома. У овакву врсту тестова спада познати Big Y-700 тест компаније FamilyTreeDNA [10].
3. Секвенцирање целог генома
Цена тестова којима се утврђује ДНК структура комплетног људског генома (енг. Whole Genome Sequencing), то јест врши секвенцирање свих 23 пара хромозома и митохондријске ДНК постаје све приступачнија. Неке компаније гарантују стопроцентну покривеност генома тестом, мада се у пракси дешава да она буде нешто мања.
II. Претходно утврђена структура ДНК може се даље анализирати и обрађивати у генеалошке сврхе:
1. Генетичка анализа аутозомних хромозома
Ова врста анализе испитује генетичку сличност две или више особа на основу 22 пара хомологних аутозомних хромозома. Сродност тестираних појединаца може се поредити по свим њиховим предачким наследним линијама. Због претходно објашњене специфичности наслеђивања, која се огледа у чињеници да свака особа на своје потомство преноси само 50% свог случајно одабраног ДНК материјала садржаног у хромозомима, предачке линије се овом методом анализе обично могу релативно поуздано поредити до пет генерација у прошлост[11].
Анализа аутозомних хромозома може се користити за утврђивање географског порекла предака и веома је популарна међу становницима земаља Новог света (Америке, Аустралија, Нови Зеланд). Такође је често користе особе које трагају за својим ближим биолошким сродницима.
Постоје различите технике анализе аутозомних хромозома. Једна од главних метода које се у ту сврху користе јесте проналажење такозваних IBD сегмената (IBD: енг. Identical by descent – идентичан по пореклу), а применљива је за поређења сродности особа које су наследиле исти аутозомни хромозом од заједничког претка. Принцип ове методе заснива се на проналажењу подударних делова између одговарајућих аутозомних хромозома појединаца чија се генетичка сродност проверава. Ови делови се називају IBD сегментима. Током гаметогенезе, појава генетичке рекомбинације на паровима хомологних хромозома доводи до размене делова између њихових ДНК ланаца. То узрокује својеврсну фрагментацију оригиналне структуре хромозома који се даље преноси на потомство. Са сваком новом генерацијом на коју се преноси одређени аутозомни хромозом, он се додатно фрагментише генетичком рекомбинацијом. Последично, што је већа сродничка удаљеност између две особе које носе заједнички предачки аутозомни хромозом, биће све краћи IBD сегменти које оне на том хромозому деле, па се може успоставити корелација између дужине IBD сегмената и броја генерација до заједничког претка особа које су у сродству.
Друга метода анализе аутозомних хромозома заснива се на поређењу SNP маркера, односно варијација на тачно одређеним SNP локусима. У комерцијалним тестовима, попут оних који нуде 23andMe, MyHeritage, AncestryDNA или FTDNA Family Finder, анализира се неколико стотина хиљада до преко 700.000 SNP маркера[12] распоређених по свим аутозомним хромозомима. Овакви тестови омогућавају да се утврди генетичка сродност између особа, али и да се процени географско порекло предака кроз поређење са референтним популацијама. Лабораторије користе биочипове дизајниране за масовно очитавање SNP маркера (генотипизација), што омогућава брзу и поуздану анализу великог броја генетичких варијација.
2. Генетичка анализа митохондријске ДНК
Анализа митохондријске ДНК омогућава поређење сродности особа по матрилинеарном генетичком наслеђу. Митохондријска ДНК се увек преноси готово неизмењена искључиво са мајке на дете, па је путем ње могуће пратити генетичку сродност појединаца до далеке прошлости. Проблеми са овом врстом анализе су како биолошке тако и социолошке природе. Биолошки проблем лежи у учесталости појављивања мутација на митохондријској ДНК, које настају много ређе него што је то случај са мутацијама на Y хромозомима.[13] Иако се на први поглед ово чини добром карактеристиком, недостатак мутација доводи до превелике униформности митохондријске ДНК, односно до слабог гранања mtDNA филогенетичког стабла, због чега се теже долази до поузданих закључака о стварној родбинској блискости тестираних појединаца.
Социолошки проблем mtDNA анализе је то што су људска друштва од прадавних времена заснована на патријархату, те да се последично у историјским изворима углавном могу проналазити подаци за патрилинеарне наследне линије, док остале ретко сежу даље од неколико генерација. У комбинацији са слабом диверсификацијом mtDNA генетичког стабла, наслеђена социолошка ограничења чине mtDNA анализу мање употребљивом у генеалошке сврхе.
3. Генетичка анализа Y хромозома
Ова врста анализе ДНК заснива се на поређењу Y хромозома између тестираних појединаца. Како овај хромозом не поседују жене, помоћу Y-DNA анализе може се пратити само патрилинеарна родбинска веза, то јест генетичко наслеђе које се преноси искључиво са оца на сина. За традиционално родословље које прати мушке наследне линије ова врста анализа је од највећег значаја. Постоје две врсте анализа Y хромозома, од којих се једна заснива на поређењу STR маркера, док се друга заснива на проналажењу SNP варијација.
ФИЛОГЕНЕТИЧКА СТАБЛА И ХАПЛОГРУПЕ
Једном стечене SNP варијације на Y хромозому преносе се са оца на сина, чиме се у сваком човеку акумулирају промене које су настале на овом хромозому код свих његових предака по мушкој наследној линији. Пажљивим поређењем присутних SNP варијација код тестираних појединаца и применом хеуристичких и статистичких метода научници су успели да формирају Y-DNA филогенетичко стабло људског рода. Ово стабло представља својеврсни родослов мушке популације човечанства, а његово гранање одређено је присутним SNP варијацијама. У корену овог стабла налази се „Y-хромозомски Адам“, односно реконструисани Y хромозом мушкарца који је претпостављени најстарији патрилинеарни предак свих досад тестираних мушкараца. Он је према тренутним прорачунима живео пре 237.000 до 581,000 година.[14] Важно је разумети да то није био једини мушкарац времена у којем је живео, али се само његова мушка лоза наставила без прекида све до данас. С обзиром да се ради о реконструисаном моделу Y хромозома нашег најстаријег мушког претка, „титула“ Y-хромозомског Адама се са новим открићима још старијих грана Y-DNA филогенетичког стабла може померати још даље у прошлост.[15]
Са растом броја тестираних појединаца и откривањем нових SNP варијација Y-DNA филогенетичко стабло се стално даље грана и унапређује.[16] Статистичким методама се може одредити вероватноћа појављивања SNP мутација у свакој новој генерацији, па је одређеним прорачунима могуће проценити време формирања појединачних грана филогенетичког стабла.
За све појединце код којих је присутна одређена SNP варијација која формира неку грану на Y-DNA филогенетичком стаблу кажемо да припадају Y-DNA хаплогрупи која је одређена том варијацијом. Тако на пример, сви мушкарци који поседују SNP варијацију I-M170 припадају хаплогрупи I-M170, која се још назива и хаплогрупа I и формирана је пре око 43.000 година. Ова грана филогенетичког стабла даље се грана на све млађе подгране, одређене млађим SNP мутацијама, које дефинишу и млађе хаплогрупе. Свака млађа грана у себи носи све SNP варијације присутне у старијим гранама из којих проистиче. Тако на пример, припадници грана I1 и I2 носе SNP варијацију I-M170, која је карактеристична за све припаднике хаплогрупе I из које ове гране произилазе, али такође поседују и своје карактеристичне SNP варијације, специфичне само за грану I1, односно I2.
Договором је усвојена подела стабла на двадесетак главних хаплогрупа, које се означавају великим словима енглеског писма, а које се даље гранају на бројне подгране.
Попут Y-DNA филогенетичког стабла, постоји и mtDNA филогенетичко стабло, које се грана по SNP варијацијама откривеним у митохондријској ДНК и представља својеврсно матрилинеарно родословно стабло људског рода. Еквивалентно, постоје такође и mtDNA хаплогрупе. Слично „Y‑хромозомском Адаму“, у генетичкој генеалогији постоји и појам „митохондријске Еве“. Он означава најстаријег заједничког матрилинеарног претка свих данашњих људи, односно жену чија се митохондријска ДНК пренела на све живе особе путем мајчине наследне линије. Као и у случају Y‑хромозомског Адама, ради се о статистичком моделу заснованом на анализи SNP варијација у митохондријској ДНК. Митохондријска Ева није била једина жена свог времена, већ једина чија је матрилинеарна линија опстала до данас без прекида. На основу молекуларног часовника процењује се да је митохондријска Ева живела пре отприлике 150.000 до 200.000 година у Африци, што се поклапа са археогенетичким налазима и моделима о афричком пореклу савременог човека.[17]
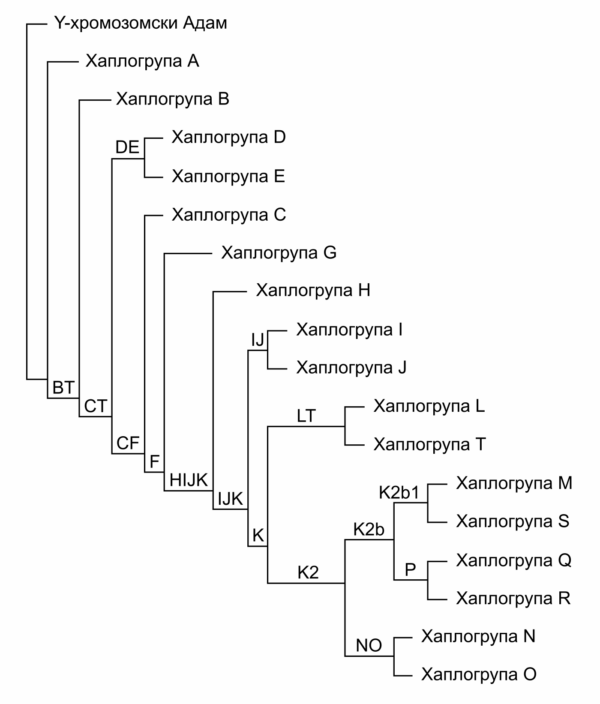 Поједностављено филогенетичко стабло Y-DNA хаплогрупа. [18]
Поједностављено филогенетичко стабло Y-DNA хаплогрупа. [18]
ОДРЕЂИВАЊЕ ПРИПАДНОСТИ Y-DNA ХАПЛОГРУПИ
За поуздано одређивање припадности некој хаплогрупи потребно је урадити Y-SNP тест којим се потврђује присуство циљане SNP варијације. С обзиром на то да постоји веома велики број могућих SNP варијација чије би се присуство могло проверавати, овакви тестови, којима се испитују свe SNP варијације на хромозому, прилично су скупи. Они спадају у помињане типове тестова којима се секвенцира одређени сегмент генома или цео геном. Могуће је и тестирање на појединачне SNP варијације, али оно представља „тражење игле у пласту сена“, што опет може резултовати великим трошковима. Да би се потрага за присутним SNP варијацијама сузила и имала прихватљиву цену, користи се друга врста Y-DNA генетичког теста – Y-STR тест. Идеја се заснива на претпоставци да је појава Y-SNP мутација праћена одговарајућим Y-STR мутацијама, те да припадници одређене хаплогрупе поседују сличне Y-STR хаплотипове. Емпиријском анализом расположивих резултата тестирања оваква претпоставка показала се тачном. Свакој хаплогрупи одговара одређени, мање или више карактеристичан хаплотип, чији се Y-STR маркери јављају код највећег броја припадника те хаплогрупе. Овакав хаплотип назива се модални хаплотип хаплогрупе, а за маркере који га формирају каже се да имају модалну вредност.
Одређивањем Y-STR хаплотипа тестиране особе може се с великом вероватноћом утврдити хаплогрупа којој та особа припада, чиме се значајно сужава број SNP маркера које је потребно додатно проверавати у циљу откривања најмлађе гране филогенетичког стабла којој припада тестирани појединац. Треба ипак нагласити да се утврђивање припадности хаплогрупи на основу Y-STR теста заснива на принципима теорије вероватноће и математичке статистике. Ту нам у помоћ долазе рачунарски програми предиктори, који на основу хаплотипа одређују највероватнију припадност хаплогрупи. Што је већи број утврђених YSTR маркера у хаплотипу, то се хаплогрупа може утврдити са већом прецизношћу и открити њена најмлађа грана. Ипак, за апсолутну потврду припадности некој хаплогрупи неопходно је урадити SNP тест.
Слично као за хаплогрупу, модални хаплотип може се одредити и за било коју групу тестираних појединаца који припадају истој грани филогенетичког стабла. Што су разлике на YSTR маркерима између тестираних појединаца мање, то су већи изгледи да они деле заједничког мушког претка у ближој прошлости. Постоје алгоритми којима се на основу сличности хаплотипова може мање или више поуздано проценити време до најмлађег заједничког претка две особе (енглески Time to most recent common ancestor, или скраћено TMRCA). Ове процене треба ипак узимати с дозом резерве, с обзиром на то да представљају само највероватнији временски распон, који у стварности може понекад и значајно да варира.
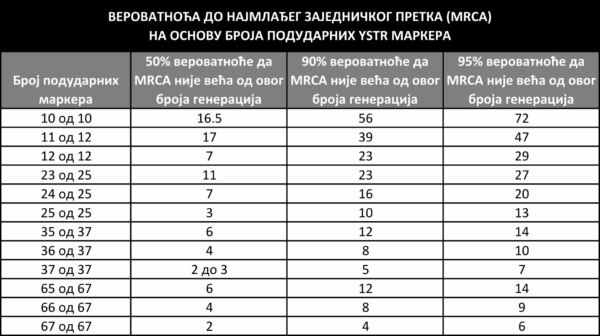
Једна процена вероватноће да особе чији се хаплотипови подударају
или су блиски деле заједничког претка у задатом броју генерација. [18.2]
Вероватноћа појаве мутације на YSTR маркерима подлеже законима који се јављају код свих случајних природних процеса: што се вредност YSTR маркера више разликује од модалне за посматрану хаплогрупу, то је мања вероватноћа да се она појави. Такође, што је одређени YSTR маркер склонији бржим променама, то је (језиком статистике речено) његово средње апсолутно одступање од модалне вредности веће. Другим речима, код хаплотипова припадника одређене хаплогрупе, на брзо променљивим маркерима ће се чешће јављати вредности различите од модалне. Може се рећи да вредности маркера које се ретко појављују представљају особеност хаплотипова у којима се јављају, па се за њих каже да представљају карактеристичне вредности маркера.
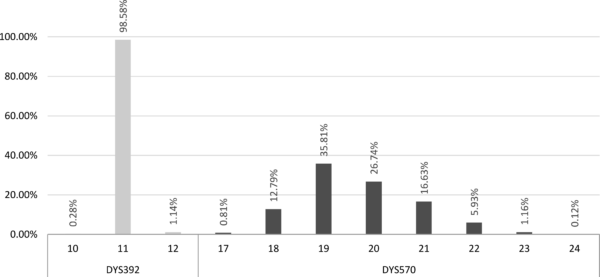
Учесталост појављивања различитих вредности
споро променљивог маркерa DYS392 и брзо променљивог DYS570 код хаплогрупе E-V13
На слици је приказан пример расподеле учесталости појављивања вредности маркера DYS392 и DYS570 на једном узорку хаплогрупе E-V13.[19] Као што се на графикону види, за споро променљиви маркер DYS392 модална вредност је 11 и јавља се у више од 98% случајева, док су вредности 10 и 12 јављају веома ретко и могу се сматрати карактеристичним. Код другог маркера, DYS570, модална вредност 19 јавља се код тек нешто више од 35% тестираних појединаца. Код овог маркера се вредност 20 не може сматрати карактеристичном, иако одступа од модалне, с обзиром на то да се прилично често појављује. Због велике варијације маркера DYS570 чак се и вредности 18 и 21 могу сматрати тек нешто ређим, док се једино за вредности 17, 22, 23 и 24 може рећи да су карактеристичне.[20]
Сличност хаплотипова тестираних појединаца код којих се јављају карактеристични маркери повећава вероватноћу да припадају истој хаплогрупи, то јест да међу њима постоји ближа генетичка веза, под условом да подударност постоји на карактеристичним маркерима. Важи и супротно: о стварној блискости појединаца који поседују модалне хаплотипове, или оне који не поседују карактеристичне маркере, незахвално је изводити закључке без додатних SNP тестова.
За групу хаплотипова која поседује карактеристичне маркере, који је јасно издвајају из основне хаплогрупе којој припадају, каже се да је добро профилисана. Добро профилисан скуп хаплотипова по правилу припада млађој грани основне хаплогрупе, коју карактеришу и одговарајуће SNP варијације. На пример, уколико хаплотипови групе тестираних појединаца који припадају хаплогрупи I2-Z17855 поседују карактеристичне маркере DYS385ab=14-14 и DYS448=21, они по правилу припадају млађој грани I2-PH3414. Ова грана је карактеристична за херцеговачки род Мириловића, те се за све хаплотипове који су блиски модалном хаплотипу хаплогрупе I2-Z17855, а поседују поменуте карактеристичне YSTR маркере, може основано претпоставити да припадају роду Мириловића. Следећи ову методологију могуће је изводити мање или више поуздане закључке о припадности хаплотипова одговарајућим гранама филогенетичког стабла, то јест генетичким родовским групама.
♦ ♦ ♦ ♦ ♦
У трећем наставку даћемо преглед основних Y‑DNA хаплогрупа у свету. Биће речи о њиховој старости, географској распрострањености и значају у историјском контексту, као и о хаплогрупама заступљеним код Срба.
[1] Не треба заборавити да је 99.9% структуре генома код свих људи идентично.
[2] Применом савремених технологија и научних сазнања процес секвенцирања ДНК временом се усавршавао и развијао, што је праћено и драстичним падом цене секвенцирања људског генома у 21. веку. С невероватних 100 милиона долара, колико су износили трошкови секвенцирања једног људског генома 2001. године, данашња цена пала је на мање од 500 долара (Видети: DNA Sequencing Costs: Data, National Human Genome Research Institute, 2023, https://www.genome.gov/about-genomics/fact-sheets/DNA-Sequencing-Costs-Data, приступљено 29.11.2025).
[3] Ово се односи на тестове којима се испитују аутозомни хромозоми, попут 23andMe, MyHeritage FamilyFinder и сличних. Ови тестови се ослањају на методу која се назива генотипизација, а којом се помоћу посебно дизајнираних биочипова истовремено очитава по неколико стотина хиљада одабраних СНП маркера.
[4] Извор: Becky Hill, John Butler, and Mike Coble, Variability of Y-STR Marker Sets in the NIST 1036 U.S. Population Samples, National Institute of Standards and Technology, Bode Technology 1st Annual Advanced DNA Technology, Mid-Atlantic Workshop, Charlottesville, VA, USA, September 19 & 20, 2012.
[5] https://worldwide.promega.com/products/forensic-dna-analysis-ce/str-amplification/powerplex-y23-system/?catNum=DC2305, приступљено 29.11.2025.
[6] https://www.qiagen.com/us/products/human-id-and-forensics/investigator-solutions/investigator-argus-y-28-qs-kit, приступљено 29.11.2025.
[7] https://www.thermofisher.com/order/catalog/product/4359513, приступљено 29.11.2025.
[8] Rhie, A., Nurk, S., Cechova, M. et al. The complete sequence of a human Y chromosome. Nature 621, 344–354 (2023). https://doi.org/10.1038/s41586-023-06457-y
[9] С обзиром да је 2023. године по први пут у потпуности секвенциран и последњи део генома који до тада није био декодиран, а који је припадао управо Y хромозому, у будућности ће се за потребе генетичке генеалогије свакако анализирати већи део Y хромозома.
[10] Big Y-700: The Forefront Of Y Chromosome Testing, 7. June 2019, https://blog.familytreedna.com/human-y-chromosome-testing-milestones/. Такође: Caleb Davis, Michael Sager, Göran Runfeldt, Elliott Greenspan, Arjan Bormans, Bennett Greenspan, and Connie Bormans, Big Y-700 White Paper. https://blog.familytreedna.com/wp-content/uploads/2019/03/big-y-700-white-paper_compressed.pdf (приступљено 29.11.2025).
[11] Autosomal DNA, https://isogg.org/wiki/Autosomal_DNA (приступљено 29.11.2025)
[12] https://support.ancestry.com/s/article/What-are-DNA-Markers?language=en_US, (приступљено 29.11.2025).
[13] Bert Ely, How do researchers trace mitochondrial DNA over centuries?, Scientific American, 2006, https://www.scientificamerican.com/article/how-do-researchers-trace/ (приступљено 29.11.2025).
[14] Видети: https://isogg.org/wiki/Y-chromosomal_Adam
[15] Године 2012. у Камеруну је откривено жариште до тада непознате прастаре Y хаплогрупе која је добила ознаку A00. Новооткривена хаплогрупа је померила време у којем је живео Y-хромозомски Адам десетинама хиљада година у прошлост, чак за више од 100.000 година. Видети: Mendez, Fernando L. et al. An African American Paternal Lineage Adds an Extremely Ancient Root to the Human Y Chromosome Phylogenetic Tree, The American Journal of Human Genetics, Volume 92, Issue 3, 454 – 459, 2013, https://doi.org/10.1016/j.ajhg.2013.02.002
[16] Како генетичке тестове и анализе спроводе различите организације и лабораторије, нажалост, не постоји јединствено стандардизовано Y-DNA филогенетичко стабло. Иако је основна структура ових стабала у свим случајевима иста, постоје разлике у виду присуства или одсуства неких млађих грана. Један од задатака организације која носи назив „Међународно удружење за генетичку генеалогију“ (International Society of Genetic Genealogy, https://isogg.org/) је да ради на стандардизацији структуре и номенклатуре Y-DNA филогенетичког стабла.
[17] ISOGG Wiki – Mitochondrial Eve, https://isogg.org/wiki/Mitochondrial_Eve (приступљено 29.11.2025).
[18] Извор: https://www.yfull.com/tree/ (приступљено 29.11.2025).
[18.2] Извор: https://www.myfamilydnatest.com/ftdna-y-37-vs-y-67-vs-y-111-vs-y-500-tests/ (приступљено 8.8.2022).
[19] Реч је о узорку сачињеном од хаплотипова хаплогрупе E-V13 који се налазе у бази Српског ДНК пројекта.
[20] Особина карактеристичности маркера не зависи, дакле, само од учесталости појављивања његове вредности унутар посматраног скупа хаплотипова, већ и од степена одступања те учесталости у односу на учесталост појављивања његове модалне вредности унутар исте групе хаплотипова.








-270x270.jpg)


















Коментари (0)