



Srpski DNK projekat je pokrenuo objavljivanje serije priloga u kojima čitaoce upoznaje sa osnovama genetičke genealogije, discipline koja poslednjih godina nailazi na sve veće interesovanje. Serijal je nastao kao dopunjena i prilagođena verzija članka „Genetika u službi rodoslovlja“ (V.Mihić, M. Rajevac, S. Jerković), objavljenog u knjizi „Genetičko poreklo Srba Stare Hercegovine“ (2021). Ovakva vrsta sadržaja do sada nije bila dostupna na srpskom jeziku u formi naučno-popularne građe onima koji su želeli da se upoznaju sa osnovama ove nove naučne oblasti. Zato smo pripremili „Malu školu genetičke genealogije“ kroz koju ćemo dati jasna objašnjenja i primere koji približavaju pojmove i metode savremene genetike, kao i njihovu primenu u istraživanju porekla porodica, rodova i naroda.
U prvom nastavku serijala predstavili smo osnovne pojmove genetike: šta su i kako funkcionišu geni i DNK, šta su hromozomi i kako se odvija reprodukcija i nasleđivanje. Bilo je reči i o ljudskom genomu, genetičkim mutacijama, kao i o STR i SNP markerima koji čine osnovu savremenih genetičkih testova.
Drugi nastavak sadrži objašnjenje vrsta i namene genetičkih testova, kao i pojam haplogrupa. Čitaoci su upoznati sa filogenetičkim stablima i metodama određivanja pripadnosti Y‑DNA haplogrupi, uz primer kako se rezultati tumače i povezuju sa rodoslovnim istraživanjima.
U trećem nastavku koji sledi dajemo pregled osnovnih Y‑DNA haplogrupa u svetu. Biće reči o njihovoj starosti, geografskoj rasprostranjenosti i značaju u istorijskom kontekstu, kao i o haplogrupama zastupljenim kod Srba.
PREGLED OSNOVNIH Y-DNA HAPLOGRUPA
Ovde ćemo dati kratak pregled svih osnovnih Y-DNA haplogrupa u svetu, zasnovan na YFull stablu i ISOGG klasifikaciji. Slika koja sledi prikazuje grananje Y-DNA haplogrupa na filogenetičkom stablu muške populacije sveta. Na grafikonu se može videti vreme izdvajanja svake od osnovnih haplogrupa, a početak širenja uspravnih linija u “trouglove” predstavlja vreme kada se procenjuje da je živeo najskoriji zajednički predak svih do sada testiranih pripadnika svake pojedinačne haplogrupe.
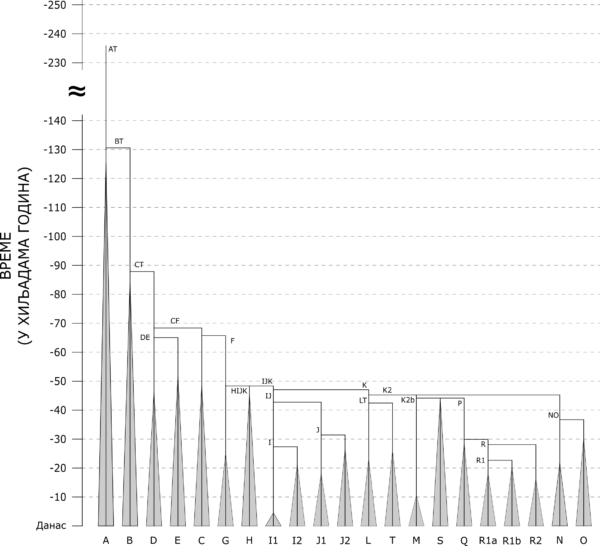
Hronologija grananja i ekspanzije osnovnih grana filogenetičkog stabla Y-DNA haplogrupa[1]
Haplogrupa A je najstarija haplogrupa, u smislu da je to predačka Y-haplogrupa celokupne današnje ljudske populacije, sa procenjenim vremenom nastanka pre oko 235.000 godina.[2] Iako se najčešće vodi kao jedinstvena haplogrupa, u stvarnosti se sastoji od čak četiri haplogrupe, redom A00, A0, A1a i A1b1, to jest sve one koje ne potpadaju pod makrohaplogrupu BT, od koje potiče preko 90% današnjih muških linija i koja je nastala od haplogrupe A1b paralelno sa sestrinskom haplogrupom A1b1. Prostorno su sve četiri A haplogrupe skoro u potpunosti ograničene na Afriku, s tim što se pojedine grane A1a i A1b1 sreću i na području Bliskog istoka i Evrope. Najveći procenti zabeleženi su među kojsanskim narodima Južne Afrike, Bušmanima i Hotentotima,[3] kao i među govornicima nilo-saharskih jezika, poput naroda Dinka i Šiluki iz Južnog Sudana[4].
Haplogrupa V izdvojila se iz makrohaplogrupe VT pre oko 88.000 godina.[5] Kao i haplogrupa A, skoro u potpunosti je ograničena na afrički kontinent, s tim što se u malom procentu sreće i na području Bliskog istoka i centralne Azije (Arabijsko poluostrvo, Iran i Avganistan). Najveće procente beleži u pojedinim predelima centralne i zapadne Afrike, pre svega među pigmejskim narodima.[6]
Haplogrupa D nastala je pre oko 65.000 godina u Africi, izdvajanjem iz haplogrupe DE.[7] Jedna je od haplogrupa koje su bile deo prvih ljudskih populacija koje su napustile afrički kontinent i započele naseljavanje Azije i Evrope. Deli se na haplogrupe D1 i D2. Grana D2 je veoma retka, zasad je pronađena samo kod nekoliko pojedinaca iz Afrike (Nigerija) i sa Bliskog istoka (Sirija i Saudijska Arabija).[8] Najbrojnija i najraširenija grana je D1, čije su podgrane prilično jasno razdeljene u nekoliko regiona. Najzastupljenija je kod starosedelačkih naroda Ainu i Rjukjuanaca,[9][10] na Tibetu[11], te kod aboridžinskih plemena Andamanskih ostrva u Indijskom okeanu.[12]
Haplogrupa E nastala je pre oko 65.000 godina u Africi[13], gde i dan danas beleži najviše procente. Ima veliki broj starih i široko rasprostranjenih grana od kojih su najzastupljenije E-M2 u zapadnoj, centralnoj i južnoj Africi među govornicima niger-kongoanskih jezika (Bantu), E-M81 na severu Afrike kod berberskih naroda, E-M123 na Bliskom istoku i E-M78 na istoku Afrike kod kušitskih naroda. Ogranak haplogrupe E-M78 je i izrazito evropska haplogrupa E-V13, koja je najprisutnija u jugoistočnoj Evropi.
Haplogrupa C nastala je pre oko 66.000 godina iz haplogrupe CF.[14] Uz haplogrupe D i F bila je deo najranijih ljudskih populacija koje su napustile afrički kontinent, i pronađena je u nekim od do danas najstarijih testiranih ljudskih skeleta.[15] Iako se ne može pohvaliti velikom brojnošću, jedna je od najraširenijih haplogrupa u svetskim okvirima, čije grane se sreću od australijskih Aboridžina[16] i Papuanaca[17], preko Indijskog potkontinenta, Kine, Japana, Sibira i centralne Azije, pa sve do Bliskog istoka i Evrope. Haplogrupa C je karakteristična haplogrupa mongolskih naroda[18].
Makrohaplogrupa F je, kao i haplogrupa C, proistekla iz CF pre oko 66.000 godina.[19] Makro u nazivu nosi zbog činjenice da je iz nje nastao veliki broj danas najbrojnijih i najraširenijih haplogrupa, i ako je posmatramo kao celinu, ona je ubedljivo najbrojnija haplogrupa u svetskim okvirima.
Haplogrupa G nastala je pre oko 48.000 godina[20] i proistekla je iz makrohaplogrupe F, najverovatnije negde na području Bliskog istoka. Karakteristična je za populacije jugozapadne Azije, pre svega Kavkaza i Anadolije, mada se javlja i u području Levanta, Iraka i Irana, i dalje na istok sve do centralne Azije.[21] U Evropi se sporadično javlja u niskim procentima od 2-5%, a najveće prisustvo beleži u regionu Alpa (10-15%) i na Sardiniji (15-30%).[22] Može se povezati s dolaskom zemljoradnje s područja Mesopotamije i Anadolije u Evropu i na Kavkaz, jer je bila ubedljivo najbrojnija haplogrupa među do sada testiranim skeletima neolitskih zemljoradnika.[23]
Haplogrupa H nastala je pre oko 48.500 godina[24], najverovatnije negde na prostoru jugozapadne Azije ili Irana. Najveći procenat današnjih nosilaca ove haplogrupe pripada grani H1, koja je skoncentrisana pre svega na Indijskom potkontinentu, gde beleži i najveće procente među dravidskim populacijama južne Indije. U Evropi haplogrupa H1 je dominantna kod romske populacije[25]. Grana H2 je danas izuzetno retka, međutim u prošlosti je bila prilično zastupljena kod neolitskih populacija Levanta, Anadolije i Evrope.[26]
Haplogrupa I nastala je pre oko 43.000 godina iz haplogrupe IJ.[27] Danas je jedna od najmnogobrojnijih i najraširenijih haplogrupa na evropskom kontinentu i verovatno najstarija haplogrupa nastala u Evropi. Pronađena je u skeletima nosilaca gravetijenske kulture, jedne od najranijih skupina ljudskih populacija koje su naseljavale Evropu u periodu paleolita.[28] Deli se na dve osnovne haplogrupe, I1 i I2. I2 danas postiže najveću koncentraciju u jugoistočnoj Evropi, posebno kod Srba, Hrvata i Bošnjaka, dok je I1 dominantna u severnoj Evropi (Skandinavija).
Haplogrupa J nastala je pre oko 43.000 godina iz haplogrupe IJ[29], najverovatnije negde na prostoru koji obuhvata istočnu Anadoliju, južni Kavkaz, zapadni Iran i severnu Mesopotamiju. Deli se na dve osnovne haplogrupe, J1 i J2. Haplogrupa J1 visoke procente beleži među semitskim narodima, Arapima i Jevrejima, pre svega na Arabijskom poluostrvu i Levantu, ali i širom Bliskog istoka i severne i severoistočne Afrike.[30] Drugo područje visoke koncentracije ove haplogrupe je Kavkaz, gde su najviši procenti zabeleženi kod severoistočnih kavkaskih naroda.[31] U Evropi su najviši procenti ove haplogrupe zabeleženi kod ostrvskih populacija Krita i Malte.[32] Haplogrupa J2 karakteristična je za današnje populacije Anadolije, Kavkaza i Mediterana. Njeno prisustvo u Evropi često se povezuje sa širenjem zemljoradnje tokom neolita. Deli se na dve osnovne grane, J2a i J2b.
Makrohaplogrupa K nastala je pre oko 47.200 godina[33], a iz nje je prostekao veliki broj nekih od danas najbrojnijih i najraširenijih haplogrupa. Iz njene grane K1 proistekle su haplogrupe L i T, iz grane K-M2335 haplogrupe N i O, a iz grane K2b haplogrupe M, S, Q i R.[34]
Haplogrupa L nastala je pre oko 42.600 godina[35] izdvajanjem iz haplogrupe LT, najverovatnije negde na prostoru jugozapadne ili centralne Azije. Deli se na dve osnovne grane, L1 i L2. Grana L2 je veoma retka i pronađena je kod svega nekolicine pojedinaca u svetu. Grana L1 dalje se deli na L1b i L1a.[36] Najveća koncentracija haplogrupe L zabeležena je u južnoj i centralnoj Aziji, pre svega u Šri Lanki, Pakistanu, Avganistanu, Indiji i Iranu. U navedenim regionima radi se skoro u potpunosti o grani L1a.[37] U Evropi, na Kavkazu i na Bliskom istoku najčešća grana haplogrupe L je L1b.[38]
Haplogrupa T je sestrinska haplogrupi L jer je takođe proistekla iz haplogrupe LT pre oko 42.600 godina.[39] Iako je raširena na veoma velikom prostoru, od zapadne Evrope do Roga Afrike i Indije, prilično je retka. Visoke procente beleži jedino kod pojedinih afričkih naroda.[40] Pronađena je u skeletima neolitskih zemljoradnika s područja Levanta i Evrope.[41] U Evropi je prisutna u niskim procentima, ali se javlja u arheogenetskim nalazima neolitskih zemljoradnika.
Haplogrupa M nastala je pre oko 44.300 godina izdvajanjem iz haplogrupe K2b1.[42] Ograničena je skoro u potpunosti na Papuu Novu Gvineju, gde beleži i najviše procente među domorodačkim populacijama, dok su manji procenti zabeleženi i među populacijama okolnih ostrva Melanezije i Indonezije[43] i među australijskim Aboridžinima.[44] Primer je „lokalne“ haplogrupe, ograničene na jedan geografski region.
Haplogrupa S nastala je kao i haplogrupa M, iz haplogrupe K2b1 pre oko 44.300 godina.[45] Takođe je najvećim delom ograničena na domorodačke populacije Papue Nove Gvineje[46], ali se javlja u značajnim procentima i kod pojedinih populacija Okeanije i jugoistočne Azije.[47]
Makrohaplogrupa P proistekla je iz makrohaplogrupe K2b pre oko 44.300 godina[48], a iz nje su nastale haplogrupe Q i R. Pronađena je u paleolitskim skeletima iz severnog Sibira starim preko 30.000 godina, koji su, moguće, pripadali predačkoj populaciji od koje potiče značajan deo današnjih populacija Sibira, centralne Azije i Evrope.[49]
Haplogrupa Q nastala je izdvajanjem iz haplogrupe P pre oko 30.000 godina[50], najverovatnije negde u južnom ili centralnom Sibiru. Zastupljena je pretežno kod sibirskih i centralnoazijskih populacija[51] i ubedljivo je najzastupljenija haplogrupa američkih Indijanaca.[52]
Haplogrupa R nastala je iz makrohaplogrupe P pre oko 30.000 godina, najverovatnije na prostoru Sibira ili centralne Azije.[53] Haplogrupa R deli se na mlađe haplogrupe R1 i R2, a R1 se deli na još mlađe R1a i R1b. Grane R1a i R1b su danas najzastupljenije u Evropi i čine osnovu većine evropskih muških linija. Haplogrupa R1a karakteristična je za evroazijski prostor, od Evrope preko Bliskog istoka i srednje Azije, do Indijskog potkontinenta i Sibira. Grana R1a-M417, kojoj pripada ogromna većina svih današnjih nosilaca R1a, po trenutno dostupnim rezultatima odigrala je jednu od ključnih uloga u nastanku i širenju indoevropskih naroda.[54] Haplogrupa R1b nastala je kada i sestrinska haplogrupa R1a, pre oko 23.000 godina[55], takođe negde na prostoru južnog Sibira ili centralne Azije. Vrlo rano je započela širenje na zapad, jer se u Evropi sreće već kod jednog skeleta iz perioda paleolita.[56] Deli se na veliki broj mlađih grana, od kojih je danas ubedljivo najbrojnija i najraširenija grana R1b-M269, koja se uz gorepomenutu granu R1a-M417 smatra ključnom u nastanku i širenju najranijih indoevropskih plemena. Od ostalih grana treba pomenuti R1b-V2219, granu koja je pronađena u mezolitskim skeletima kulture Lepenskog vira[57], a koja se danas u Evropi može pronaći samo u tragovima, dok, iznenađujuće, najviše procente beleži kod pojedinih plemena s područja Kameruna i Nigerije (preko 90%)[58], koji su najverovatnije posledica neolitske seobe iz Evrope preko severne Afrike.[59] Haplogrupa R2 nastala je pre oko 28.000 godina izdvajanjem iz haplogrupe R.[60] Pretežno je skoncentrisana na južnu i centralnu Aziju, a najveći procenat zabeležen je kod naroda Burušo u severnom Pakistanu.[61]
Haplogrupa N nastala je iz haplogrupe NO pre oko 37.000 godina[62], najverovatnije negde na prostoru (jugo)istočne Azije, gde pokazuje najveću raznovrsnost haplotipova.[63] Najviše procente beleži na prostoru severne Evroazije, dok se u manjem procentu javlja i u istočnoj i centralnoj Aziji, i na Balkanu.[64] Deli se na dve osnovne mlađe grane, N1 i N2. Grana N1 je ubedljivo brojnija i raširenija grana, koja se sreće od jugoistočne Azije, preko Sibira, sve do severne i severoistočne Evrope. Ključna je za uralske narode i često se povezuje sa širenjem uralskih jezika. Na prostoru Evroazije najveće procente beleži kod pripadnika uralske (Finaca, Hantija, Mansija, Neneca i Nganasana) i turske jezičke familije (Jakuti, Dolgani, Kakasi, Tuvanci, Tatari i Čuvaši), kao i kod Burjata, Evena, Evenka i Jukagira.[65] Grana N2 je veoma retka u svetskim okvirima, prisutna je u malom procentu u Evropi, a najveći procenat dostiže upravo kod Srba (oko 3%).[66]
Haplogrupa O nastala je takođe iz haplogrupe NO pre oko 37.000 godina, na području istočne ili jugoistočne Azije, gde je i danas najzastupljenija.[67] U manjem procentu je zastupljena i u Okeaniji, južnoj Aziji i na Madagaskaru. Deli se na dve mlađe grane, O1 i O2. Grana O1 najviše procente beleži među stanovnicima Japana[68] i Južne Koreje[69], i među narodima austroazijske[70] (jugoistočna Azija, istočna Indija i Bangladeš) i austronezijske jezičke familije[71] (Tajvan, Madagaskar, Indonezija, Filipini i Okeanija). Grana O2 je najzastupljenija kod Han Kineza (preko 50%) i njima srodnih naroda iz tibetansko-burmanske jezičke familije s područja Tibeta, Nepala i Burme, ali i kod mnogih okolnih naroda, Mandžuraca, Mongola, Korejaca, Vijetnamaca i drugih.[72] To je najbrojnija haplogrupa na svetu po broju nosilaca.
* * * * *
Sve osnovne haplogrupe se dalje se granaju na ogroman broj mlađih grana i podgrana formirajući Y-DNA filogenetičko stablo čovečanstva. Otkrivanjem novih SNP varijacija iz kojih nastaju nove haplogrupe, ovo stablo se svakog dana dopunjuje i proširuje. U novembru 2025. godine na Y-DNA filogenetičkom stablu kompanije YFull bilo je 64.704 grana, odnosno haplogrupa, koje su bile definisane sa 429.611 SNP varijacija.[73] U isto vreme Y-DNA stablo koje održava kompanije Family Tree DNA sadržalo je preko 98.000 grana (haplogrupa) i više od 837.000 SNP varijacija.[74]
ZAKLJUČCI
Muške populacije većine svetskih naroda, pogotovo evropskih, sastoje se od velikog broja različitih haplogrupa, koje su zastupljene u većem ili manjem procentu. Haplogrupe se ne mogu povezati s nacijama, jer je pripadnost određenoj haplogrupi biološka (nepromenljiva) kategorija, dok su narodi i nacije društvene (i samim tim promenljive) kategorije.
Primena metoda genetičke genealogije za istraživanje porekla porodica i naroda moguća je samo ukoliko postoji razmena genetičkih rezultata i ako se oni međusobno upoređuju i povezuju. Zato su se već od početka komercijalnih testiranja počele formirati baze genetičkih rezultata, rodoslovni ili nacionalni projekti. Pri tome je kvalitet saznanja koje takve baze daju u direktnoj vezi s veličinom baze. Svaki testirani pojedinac svojim rezultatom doprinosi upotpunjavanju i sagledavanju celokupne genetičke slike pojedinih regiona, naroda, rodova. Kako bi se u svetlu genetičkih rezultata bolje sagledala etnička prošlost srpskog naroda, formiran je nacionalni Srpski DNK projekat, s ciljem da na jednom mestu sakupi genetičke rezultate Srba, ali i naroda s kojima su Srbi u prošlosti delili prostor i istoriju. Počevši sa skromnih oko dve stotine testiranih u 2012. godini, kada je osnovan, Srpski DNK projekat je do danas izrastao u jedan od najvećih nacionalnih genetičkih projekata u Evropi, koji u svojoj bazi trenutno sadrži genetičke rezultate preko 11.000 testiranih pojedinaca.[75] Postojanje takve baze omogućava da se rezultati DNK testiranja sagledaju i obrade na ispravan način i smeste ne samo u nacionalni, srpski, već i u evropski i svetski genetički kontekst. Mogućnosti koje pruža baza Srpskog DNK projekta pomažu da se testirani pojedinci iz regionalnih istraživanja grupišu u rodove po svom patrilinearnom poreklu, kao i da se ti rodovi sigurnije profilišu.
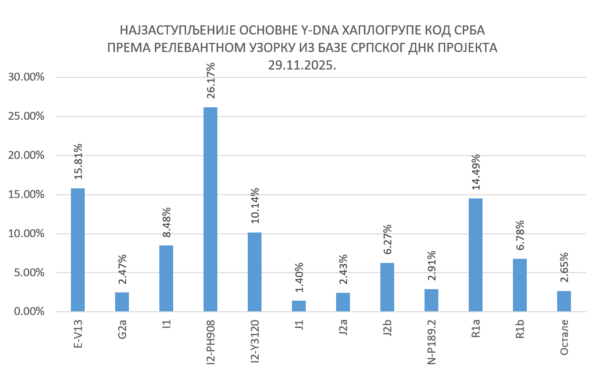
Za kraj, navodimo presek zastupljenosti Y-DNA haplogrupa kod Srba na osnovu podataka iz baze Srpskog DNK projekta:
[1] Podaci o starosti grana i vremenu življenja najmlađeg zajedničkog pretka (TMRCA) preuzeti sa https://www.yfull.com/tree/A0-T/ (pristupljeno 29. 11. 2025).
[2] https://www.yfull.com/tree/A0-T/
[3] King TE, Parkin EJ, Swinfield G, et al. (March 2007). “Africans in Yorkshire? The deepest-rooting clade of the Y phylogeny within an English genealogy”. Eur. J. Hum. Genet. 15 (3): 288–93.
[4] Hassan HY, Underhill PA, Cavalli-Sforza LL, Ibrahim ME. Y-chromosome variation among Sudanese: restricted gene flow, concordance with language, geography, and history. Am J Phys Anthropol. 2008 Nov;137(3):316-23. doi: 10.1002/ajpa.20876. PMID: 18618658.
[5] https://www.yfull.com/tree/B/
[6] Underhill PA, Shen P, Lin AA, et al. (November 2000). “Y chromosome sequence variation and the history of human populations”. Nat. Genet. 26 (3): 358–61.
[7] https://www.yfull.com/tree/D/
[8] Haber; et al. (2019). “A Rare Deep-Rooting D0 African Y-Chromosomal Haplogroup and Its Implications for the Expansion of Modern Humans out of Africa”. Genetics: genetics.302368.2019.
[9] Hammer MF, Karafet TM, Park H, Omoto K, Harihara S, Stoneking M, Horai S (2006). “Dual origins of the Japanese: common ground for hunter-gatherer and farmer Y chromosomes”. Journal of Human Genetics. 51 (1): 47–58
[10] Tajima A, Hayami M, Tokunaga K, Juji T, Matsuo M, Marzuki S, Omoto K, Horai S (2004). “Genetic origins of the Ainu inferred from combined DNA analyses of maternal and paternal lineages”. Journal of Human Genetics. 49 (4): 187–93
[11] Hua Zhong, Hong Shi, Xue-Bin Qi, Zi-Yuan Duan, Ping-Ping Tan, Li Jin, Bing Su, and Runlin Z. Ma (2011), “Extended Y Chromosome Investigation Suggests Postglacial Migrations of Modern Humans into East Asia via the Northern Route.” Mol. Biol. Evol.28(1):717–727.
[12] Shi H, Zhong H, Peng Y, Dong YL, Qi XB, Zhang F, Liu LF, Tan SJ, Ma RZ, Xiao CJ, Wells RS, Jin L, Su B (October 2008). “Y chromosome evidence of earliest modern human settlement in East Asia and multiple origins of Tibetan and Japanese populations”. BMC Biology. 6: 45.
[13] https://www.yfull.com/tree/E/
[14] https://www.yfull.com/tree/C/
[15] Fu; et al. (2016).”The genetic history of Ice Age Europe”. 10.1038/nature17993; Takođe: Sikora; et al. (2017).”Ancient genomes show social and reproductive behavior of early Upper Paleolithic foragers”. 10.1126/science.aao1807.
[16] Nagle, N.; et al. (2015). “Antiquity and diversity of aboriginal Australian Y-chromosomes”. American Journal of Physical Anthropology. 159 (3): 367–81.
[17] ISOGG, 2015 “Y-DNA Haplogroup C and its Subclades – 2015”
[18] Malyarchuk B, Derenko M, Denisova G, Wozniak M, Grzybowski T, Dambueva I, Zakharov I. Phylogeography of the Y-chromosome haplogroup C in northern Eurasia. Ann Hum Genet. 2010 Nov;74(6):539-46. doi: 10.1111/j.1469-1809.2010.00601.x. Epub 2010 Aug 20. PMID: 20726964.
[19] https://www.yfull.com/tree/F/
[20] https://www.yfull.com/tree/G/
[21] Rootsi S, Myres NM, Lin AA, et al. (December 2012). “Distinguishing the co-ancestries of haplogroup G Y-chromosomes in the populations of Europe and the Caucasus”. Eur. J. Hum. Genet. 20 (12): 1275–82.
[22] Isto
[23] Mathieson; et al. (2015).”Genome-wide patterns of selection in 230 ancient Eurasians”. 10.1038/nature16152
[24] https://www.yfull.com/tree/H/
[25] Martínez-Cruz, B., Mendizabal, I., Harmant, C. et al. Origins, admixture and founder lineages in European Roma. Eur J Hum Genet 24, 937–943 (2016). https://doi.org/10.1038/ejhg.2015.201
[26] Mathieson; et al. (2015).”Genome-wide patterns of selection in 230 ancient Eurasians”. 10.1038/nature16152; Takođe: Lazaridis; et al. (2016).”Genomic insights into the origin of farming in the ancient Near East”. 10.1038/nature19310; Lipson; et al. (2017).”Parallel palaeogenomic transects reveal complex genetic history of early European farmers”. 10.1038/nature24476.
[27] https://www.yfull.com/tree/I/
[28] Fu; et al. (2016).”The genetic history of Ice Age Europe”. 10.1038/nature17993
[29] https://www.yfull.com/tree/J/
[30] Chiaroni, Jacques; King, Roy J; Myres, Natalie M; Henn, Brenna M; Ducourneau, Axel; Mitchell, Michael J; Boetsch, Gilles; Sheikha, Issa; et al. (2010). “The emergence of Y-chromosome haplogroup J1e among Arabic-speaking populations”. European Journal of Human Genetics. 18 (3): 348–353
[31] Balanovsky, O.; Dibirova, K.; Dybo, A.; Mudrak, O.; Frolova, S.; Pocheshkhova, E.; Haber, M.; Platt, D.; et al. (2011). “Parallel evolution of genes and languages in the Caucasus region”. Molecular Biology and Evolution. 28 (10): 2905–20
[32] El-Sibai, Mirvat; Platt, Daniel E.; Haber, Marc; Xue, Yali; Youhanna, Sonia C.; Wells, R. Spencer; Izaabel, Hassan; Sanyoura, May F.; et al. (2009). “Geographical structure of the Y-chromosomal genetic landscape of the Levant: a coastal-inland contrast”. Annals of Human Genetics. 73 (Pt 6): 568–581; Takođe: King, R. J.; Özcan, S. S.; Carter, T.; Kalfoğlu, E.; Atasoy, S.; Triantaphyllidis, C.; Kouvatsi, A.; Lin, A. A.; et al. (2008). “Differential Y-chromosome Anatolian influences on the Greek and Cretan Neolithic”. Annals of Human Genetics. 72 (Pt 2): 205–214.
[33] https://www.yfull.com/tree/K/
[34] Isto
[35] https://www.yfull.com/tree/L/
[36] Sengupta, S.; Zhivotovsky, L. A.; King, R.; Mehdi, S. Q.; Edmonds, C. A.; Chow, C-E. T.; Lin, A. A.; Mitra, M.; Sil, S. K.; Ramesh, A.; Usha Rani, M. V.; Thakur, C. M.; Cavalli-Sforza, L. L.; Majumder, P. P.; Underhill, P. A. (2006). “Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists”. The American Journal of Human Genetics. 78 (2): 202–21.
[37] Di Cristofaro, Julie; Pennarun, Erwan; Mazières, Stéphane; Myres, Natalie M.; Lin, Alice A.; Temori, Shah Aga; Metspalu, Mait; Metspalu, Ene; Witzel, Michael; King, Roy J.; Underhill, Peter A.; Villems, Richard; Chiaroni, Jacques (2013). “Afghan Hindu Kush: Where Eurasian Sub-Continent Gene Flows Converge”. PLOS ONE. 8 (10): e76748. Takođe: Cordaux, Richard; Aunger, Robert; Bentley, Gillian; Nasidze, Ivane; Sirajuddin, S. M.; Stoneking, Mark (3 February 2004). “Independent origins of Indian caste and tribal paternal lineages”. Current Biology. 14 (3): 231–235; Lacau, Harlette; Gayden, Tenzin; Regueiro, Maria; Chennakrishnaiah, Shilpa; Bukhari, Areej; Underhill, Peter A; Garcia-Bertrand, Ralph L; Herrera, Rene J (18 April 2012). “Afghanistan from a Y-chromosome perspective”. European Journal of Human Genetics. 20 (10): 1063–1070.
[38] Vincenza Battaglia, Simona Fornarino, Nadia Al-Zahery, et al. (2009), “Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe.” European Journal of Human Genetics (2009) 17, 820–830; Takođe: Oleg Balanovsky, Khadizhat Dibirova, Anna Dybo, et al. (2011), “Parallel Evolution of Genes and Languages in the Caucasus Region.” Molecular Biology and Evolution 2011 October ; 28(10): 2905–2920; Bayazit Yunusbayev, Mait Metspalu, Mari Järve, et al. (2012), “The Caucasus as an Asymmetric Semipermeable Barrier to Ancient Human Migrations.” Molecular Biology and Evolution 29(1):359–365.
[39] https://www.yfull.com/tree/T/
[40] Mendez FL, Karafet TM, Krahn T, Ostrer H, Soodyall H, Hammer MF (2011). “Increased resolution of Y chromosome haplogroup T defines relationships among populations of the Near East, Europe, and Africa”. Human Biology. 83 (1): 39–53.
[41] Lazaridis; et al. (2016).”Genomic insights into the origin of farming in the ancient Near East”. Nature, nature19310; Takođe: Mathieson; et al. (2015).”Genome-wide patterns of selection in 230 ancient Eurasians”. Nature, volume528, 499–503.
[42] https://www.yfull.com/tree/M/
[43] Kayser, Manfred; Brauer, Silke; Weiss, Gunter; Schiefenhövel, Wulf; Underhill, Peter; Shen, Peidong; Oefner, Peter; Tommaseo-Ponzetta, Mila; Stoneking, Mark (2003). “Reduced Y-Chromosome, but Not Mitochondrial DNA, Diversity in Human Populations from West New Guinea”. The American Journal of Human Genetics. 72 (2): 281–302. Takođe: Karafet, T. M.; Mendez, F. L.; Meilerman, M. B.; Underhill, P. A.; Zegura, S. L.; Hammer, M. F. (2008). “New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree”. Genome Research. 18 (5): 830–8.
[44] Nagle, N.; et al. (2015). “Antiquity and diversity of aboriginal Australian Y-chromosomes”. American Journal of Physical Anthropology. 159 (3): 367–81.
[45] https://www.yfull.com/tree/S/
[46] Kayser, Manfred; Brauer, Silke; Weiss, Gunter; Schiefenhövel, Wulf; Underhill, Peter; Shen, Peidong; Oefner, Peter; Tommaseo-Ponzetta, Mila; Stoneking, Mark (2003). “Reduced Y-Chromosome, but Not Mitochondrial DNA, Diversity in Human Populations from West New Guinea”. The American Journal of Human Genetics. 72 (2): 281–302.
[47] Scheinfeldt, L.; Friedlaender, F; Friedlaender, J; Latham, K; Koki, G; Karafet, T; Hammer, M; Lorenz, J (2006). “Unexpected NRY Chromosome Variation in Northern Island Melanesia”. Molecular Biology and Evolution. 23 (8): 1628–41. Takođe: Cox, Murray P.; Mirazón Lahr, Marta (2006). “Y-chromosome diversity is inversely associated with language affiliation in paired Austronesian- and Papuan-speaking communities from Solomon Islands”. American Journal of Human Biology. 18 (1): 35–50.
[48] https://www.yfull.com/tree/P/
[49] Sikora; et al. (2019). “The population history of northeastern Siberia since the Pleistocene”. Nature 570, 182–188
[50] https://www.yfull.com/tree/Q/
[51] Jeffrey, T.; et al. (Jan 2002). “The Dual Origin and Siberian Affinities of Native American Y Chromosomes”. Am J Hum Genet. 70 (1): 192–206. Takođe: Malyarchuk, Boris; et al. (2011). “Ancient links between Siberians and Native Americans revealed by subtyping the Y chromosome haplogroup Q1a”. Journal of Human Genetics. 56 (8): 583–588; Dulik; et al. (2012).” Mitochondrial DNA and Y Chromosome Variation Provides Evidence for a Recent Common Ancestry between Native Americans and Indigenous Altaians”. Am J Hum Genet., 90(2): 229–246.
[52] Zegura, S. L.; Karafet, TM; Zhivotovsky, LA; Hammer, MF (2004). “High-Resolution SNPs and Microsatellite Haplotypes Point to a Single, Recent Entry of Native American Y Chromosomes into the Americas” (PDF). Molecular Biology and Evolution. 21 (1): 164–75. Takođe: Malhi, RS; Gonzalez-Oliver, A; Schroeder, KB; Kemp, BM; Greenberg, JA; Dobrowski, SZ; Smith, DG; Resendez, A; Karafet, T; Hammer, M; Zegura, S; Brovko, T (2008). “Distribution of Y chromosomes among native North Americans: a study of Athapaskan population history”. Am. J. Phys. Anthropol. 137 (4): 412–24.
[53] https://www.yfull.com/tree/R/
[54] Allentoft; et al. (2015). “Population genomics of Bronze Age Eurasia”. Nature, volume 522, 167–172. Takođe: Mittnik; et al. (2018).”The genetic prehistory of the Baltic Sea region”. Nature Communications volume 9, Article number: 442.
[55] https://www.yfull.com/tree/R1b/
[56] Fu; et al. (2016). “The genetic history of Ice Age Europe”. 10.1038/nature17993
[57] Mathieson; et al. (2018). “The genomic history of southeastern Europe”. 10.1038/nature25778
[58] Wood, ET; Stover, DA; Ehret, C; Destro-Bisol, G; Spedini, G; Mcleod, H; Louie, L; Bamshad, M; et al. (2005). “Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes” (PDF). European Journal of Human Genetics. 13 (7): 867–76.
[59] D’Atanasio; et al. (2018). “The peopling of the last Green Sahara revealed by high-coverage resequencing of trans-Saharan patrilineages”. Genome Biol 19, 20
[60] https://www.yfull.com/tree/R2/
[61] Di Cristofaro, Julie; Pennarun, Erwan; Mazières, Stéphane; Myres, Natalie M.; Lin, Alice A.; Temori, Shah Aga; Metspalu, Mait; Metspalu, Ene; Witzel, Michael; King, Roy J.; Underhill, Peter A.; Villems, Richard; Chiaroni, Jacques (2013). “Afghan Hindu Kush: Where Eurasian Sub-Continent Gene Flows Converge”. PLOS ONE. 8 (10): e76748.
[62] https://www.yfull.com/tree/N/
[63] Rootsi, Siiri; Zhivotovsky, Lev A; Baldovič, Marian; Kayser, Manfred; Kutuev, Ildus A; Khusainova, Rita; Bermisheva, Marina A; Gubina, Marina; et al. (2006). “A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe”. European Journal of Human Genetics. 15 (2): 204–11.
[64] Ilumäe; et al. (2016). “Human Y Chromosome Haplogroup N: A Non-trivial Time-Resolved Phylogeography that Cuts across Language Families”. American Journal of Human Genetics. 99 (1): 163–73.
[65] Isto
[66] Srpski DNK projekat; https://dnk.poreklo.rs/DNK-projekat/
[67] https://www.yfull.com/tree/O/
[68] Hammer, Michael F.; Karafet, Tatiana M.; Park, Hwayong; Omoto, Keiichi; Harihara, Shinji; Stoneking, Mark; Horai, Satoshi (2005). “Dual origins of the Japanese: Common ground for hunter-gatherer and farmer Y chromosomes”. Journal of Human Genetics. 51 (1): 47–58.
[69] Kim, SH; Kim, Ki-Cheol; Shin, Dong-Jik; Jin, Han-Jun; Kwak, Kyoung-Don; Han, Myun-Soo; Song, Joon-Myong; Kim, Won; Kim, Wook (2011). “High frequencies of Y-chromosome haplogroup O2b-SRY465 lineages in Korea: a genetic perspective on the peopling of Korea”. Investigative Genetics. 2 (10)
[70] Kumar; et al. (2007). “Y-chromosome evidence suggests a common paternal heritage of Austro-Asiatic populations”. BMC Evolutionary Biology 2007, 7:47; Takođe: Reddy BM, Langstieh BT, Kumar V, Nagaraja T, Reddy ANS, et al (2007) Austro-Asiatic Tribes of Northeast India Provide Hitherto Missing Genetic Link between South and Southeast Asia. PLoS ONE 2(11): e1141.
[71] Kayser, M.; Choi, Y.; Van Oven, M.; Mona, S.; Brauer, S.; Trent, R. J.; Suarkia, D.; Schiefenhovel, W.; Stoneking, M. (2008). “The Impact of the Austronesian Expansion: Evidence from mtDNA and Y Chromosome Diversity in the Admiralty Islands of Melanesia”. Molecular Biology and Evolution. 25 (7): 1362–74; Takođe: Karafet, T. M.; Hallmark, B.; Cox, M. P.; Sudoyo, H.; Downey, S.; Lansing, J. S.; Hammer, M. F. (2010). “Major East-West Division Underlies Y Chromosome Stratification across Indonesia”. Molecular Biology and Evolution. 27 (8): 1833–44.
[72] Shi, Hong; Dong, Yong-li; Wen, Bo; Xiao, Chun-Jie; Underhill, Peter A.; Shen, Pei-Dong; Chakraborty, Ranajit; Jin, Li; Su, Bing (2005). “Y-Chromosome Evidence of Southern Origin of the East Asian–Specific Haplogroup O3-M122”. The American Journal of Human Genetics. 77 (3): 408–19.
[73] https://www.yfull.com/tree/, pristupljeno 29.11.2025.
[74] https://www.familytreedna.com/public/y-dna-haplotree/, pristupljeno 29.11.2025.
[75] Videti: https://dnk.poreklo.rs/DNK-projekat/.








-270x270.jpg)


















Komentari (0)